
Nádorové mikroprostředí
Tumor Microenvironment
Cancer microenvironment plays an essential role in promoting the growth, proliferation and ability of cancer cells to establish metastatic deposits. This microenvironment consists predominantly of cancer-associated fibroblasts and the extracellular matrix produced by them, immune cells (macrophages, granulocytes, mast cells and lymphocytes), blood vessels, adipocytes and mesenchymal stem cells. These cells interact with each other, both via intercellular communication and signal molecules. The signal molecules include, for example, growth factors, chemokines, interleukins or galectins. In the review article, we present the current knowledge about individual cell groups and their interaction. The presented information relates mainly to the already applied or future prospective clinical use.
Keywords:
Stem cells – cancer-associated fibroblasts – extracellular matrix – immune cells – growth factors – chemokines – interleukins – galectin
:
Š. Novák 1,2; V. Bandurová 1,2; A. Mifková 1,2; D. Kalfeřt 1; Z. Fík 1; P. Lukeš 1; P. Szabó 2,3; J. Plzák 1; K. Smetana Jr. 2,3
:
Otorinolaryngol Foniatr, 68, 2019, No. 1, pp. 41-51.
:
Review Article
Nádorové mikroprostředí hraje esenciální roli v podpoře růstu, proliferace a schopnosti nádorových buněk zakládat metastatická ložiska. Je tvořeno převážně z nádorově asociovaných fibroblastů a jimi produkované extracelulární matrix, imunitních buněk (makrofágy, granulocyty, mastocyty a lymfocyty), cév, adipocytů a mezenchymových kmenových buněk. Tyto buňky se vzájemně ovlivňují, a to jak pomocí intercelulární komunikace, tak prostřednictvím signálních molekul. Mezi signální molekuly patří například růstové faktory, chemokiny, interleukiny nebo galektiny. V přehledovém článku prezentujeme současné poznatky o jednotlivých skupinách buněk a jejich vzájemné interakci. Prezentované informace jsou vztaženy zejména k již aplikovanému nebo v budoucnu perspektivnímu klinickému využití.
Klíčová slova:
kmenové buňky – nádorově asociované fibroblasty – extracelulární matrix – imunitní buňky – růstové faktory – chemokiny – interleukiny – galektiny
ÚVOD
Incidence nádorových onemocnění má v posledních desetiletích vzestupnou tendenci (81). Vzrůstající počet nových onemocnění je, mimo rizikové chování, způsoben životním stylem, stárnutím populace, a tím podmíněnou nestabilitou genomu u starších jedinců (111).
S přibližně 550 000 každoročně nově diagnostikovanými případy zůstávají karcinomy hlavy a krku celosvětově významnou příčinou nemocnosti a úmrtnosti (137). Mezi hlavní etiologické faktory patří kouření, konzumace alkoholu a infekce lidskými papilomaviry (HPV). Velké evropské i americké epidemiologické studie v posledních 40 letech popisují vzestupný trend v incidenci karcinomů orofaryngu, a to zejména u mladších jedinců (47, 49). Incidence karcinomů v jiných lokalitách horních cest dýchacích a polykacích má spíše sestupnou tendenci (91). Vzestup počtu nově diagnostikovaných případů (a to i u nekuřáků) je přisuzován zvyšující se prevalenci vysoce rizikových typů – HPV u mladších jedinců (112). I přes značný pokrok v diagnostice a léčbě jsou dvě třetiny karcinomů hlavy a krku diagnostikovány v pozdních stadiích onemocnění a přibližně 50 % pacientů zemře na karcinom do dvou let od stanovení diagnózy (27). Současné možnosti léčby jsou zatíženy nežádoucími účinky, a to buď mutilací po rozsáhlejších chirurgických výkonech, vedlejšími účinky radioterapie, nebo časnou i pozdní toxicitou chemoterapie.
Na vzniku nádorů se podílejí genetické a regulační změny, na jejichž základě nastává funkční deregulace dělení a přirozeného zániku buněk. Nekontrolované dělení nádorových buněk vede k jejich enormní akumulaci a ztrátě nezávislosti na okolí (119). Je evidentní, že interakce maligně transformované buněčné populace a okolní tkáně, tvořené jinými netransformovanými buněčnými typy a mezibuněčnou hmotou (nádorové stroma), je podstatným faktorem v patogenezi nádorových onemocnění. Epitelově-mezenchymové interakce zde tedy sehrávají významnou úlohu při tvorbě specifického prostředí, kde dochází k vzájemnému ovlivnění jak nádorového parenchymu, tak nádorového stromatu (36). Dřívější předpoklad, že nádorové stroma představuje pouze podpůrnou tkáň nádorů, je již překonán a v současné nádorové biologii je mu připisována významná role umožňující přímé ovlivnění biologické funkce nádorových buněk (71, 75, 98). Prognostický význam vyššího objemu stromatu (desmoplastická stroma) byl potvrzen u řady typů nádorů, včetně karcinomu prsu, tlustého střeva, plic a jícnu (22, 82, 135, 136). Stroma se skládá z nádorově asociovaných fibroblastů (cancer associated fibroblasts=CAF) a jimi produkované extracelulární matrix (ECM), imunitních buněk (makrofágy, granulocyty, mastocyty a lymfocyty), cév, adipocytů a mezenchymových kmenových buněk (48, 83, 100) (obr. 1).

KMENOVÉ BUŇKY A NÁDORY
Kmenové buňky u dospělého organismu jsou málo diferencované, převážně monopotentní a multipotentní buňky, nacházející se ve většině lidských tkání. Jejich základní funkcí je tkáňová sebeobnova. Slouží i k obnově poškozené nebo opotřebované tkáně a udržují homeostázu organismu. Za jistých okolností je ovšem možná jejich přeměna v maligní kmenové buňky (cancer stem cells=CSC). Existence CSC byla nejdříve popsána u hematologických malignit, avšak jejich přítomnost byla později prokázána i u solidních tumorů (48). Například epidermové kmenové buňky (ESC) představují jeden ze zdrojů nádorových buněk dlaždicového karcinomu (spinocellular cancer cells=SCC) (96). Pro zachování málo diferencovaného fenotypu kmenových buněk, včetně nádorových, je velmi důležité okolní mikroprostředí nazvané niche (134). Toto mikroprostředí s jeho specifickými cytokiny může být (například u chronického zánětu) spouštěčem maligního reprogramování CSC (114). Vzhledem k pomalému proliferačnímu cyklu CSC a jejich schopnosti vyloučit z cytoplazmy xenobiotika, je velmi obtížné tyto buňky účinně terapeuticky ovlivnit (24). Tato vlastnost může být vysvětlením selhání nádorové chemoterapie nebo minimálního reziduálního onemocnění (89).
NÁDOROVĚ ASOCIOVANÉ FIBROBLASTY
Jednu z nejpočetnějších buněčných populací nádorového stromatu představují CAF. Morfologicky se jedná o vřetenité buňky, které jsou charakteristické expresí vimentinu, α-SMA (alpha-smooth muscle actin) a FAP (fibroblast activation protein). FAP je jedním z možných cílů protinádorové terapie. Při použití monoklonální protilátky proti FAP byla experimentálně prokázána kompletní regrese růstů nádoru plic, pankreatu a nádorů hlavy a krku (94). Další z proteinů asociovanými s CAF jsou FSP-1 (fibroblast specific protein-1), PDGFR (platelet-derived growth factor receptor), podoplanin-a, S100A4 (S100 calcium-binding protein A4) a IGFBP-7 (insulin-like growth factor-binding protein 7) (17, 64, 104). Rozdíl mezi zdravými fibroblasty (NF) a CAF lze prokázat na podkladě významné změny expresního profilu, a to až u 640 genů (u CAF z bazocelulárního karcinomu) (85).
CAF hrají velmi důležitou roli v utváření a zachování optimálního mikroprostředí pro přežití a progresi nádorových buněk, včetně kmenových (79). Je prokázána schopnost CAF transformovat fibroblasty na buňky podobné kmenovým, např. u nádoru pankreatu (46), které pak ovlivní progresi nádoru. CAF z bazocelulárního karcinomu jsou schopné indukovat expresi markerů embryonálních kmenových buněk Oct-4 a Nanog u myších fibroblastů, a tyto fibroblasty jsou dále schopné transformace v rozsahu podobném mezenchymovým kmenovým buňkám (123).
CAF mají výrazný efekt na proliferaci, epitelo-mezenchymový přechod a migraci nádorových buněk u různých typů nádorů (38, 80, 93). Také proces růstu a vytváření sekundárních nádorových ložisek je podpořen aktivitou CAF. Výjimečně však existují i nálezy, kdy byl pozorován i jejich opačný efekt, například u nádorů pankreatu. U karcinomu prsu byla přítomnost CAF asociovaná s produkcí CXCL-12 (C-X-C motif chemokine ligand 12) a IGF-1 (insulin-like growth factor 1). Tyto faktory jsou u nádoru prsu specifické pro kostní metastázy, lze proto předpokládat, že analýzou CAF lze predikovat typ metastatického postižení (149).
Fenotyp CAF se nápadně podobá aktivovaným fibroblastům v hojící se ráně (tedy myofibroblastům), které exprimují α-SMA (56) a jsou odpovědné za kontrakci hojící se rány. Tato podoba byla popsána již před 30 lety (28). Zásadní rozdíl spočívá v časově neomezené aktivaci CAF v nádorovém mikroprostředí (124). Původ CAF nebude u všech nádorů identický. Existuje nejspíš hned několik způsobů, kterými mohou nádorově asociované fibroblasty vzniknout: 1) aktivací lokálních fibroblastů; 2) epitelově-mesenchymovým přechodem (EMT) vlastních nádorových buněk; 3) diferenciací z migrujících mesenchymových kmenových buněk, pocházejících z kostní dřeně atrahovaných do místa nádoru (86). Připustit lze pravděpodobně i kombinaci mechanismů vzniku v závislosti na buněčném typu a signálech okolí (71). CAF jsou při kokultivaci se zdravými keratinocyty schopné vyvolat u epitelových buněk expresi mesenchymového znaku vimentinu, jehož společná koexprese s keratinem je jedním ze znaků epitelově-mesenchymového přechodu (126). Data z experimentů na myších modelech s transplantovanými lidskými nádorovými buňkami naznačují, že CAF vznikají především aktivací lokálního mesenchymu hostitele (29). Proces aktivace CAF z NF je způsoben jak parakrinně pomocí TGF-β (transforming growth factor β), EGF (epidermal growth factor) PDGF (platelet-derived growth factor), FGF-2 (fibroblast growth factor 2) nebo CXCL-12 (C-X-C motif chemokine ligand 12) (102), tak i přímým kontaktem buněk pomocí adhezivních molekul jako ICAM-1 (intercellular adhesion molecule 1), nebo VICAM-1 (vascular-cell adhesion molecule 1) (19). Nejnovější způsob aktivace CAF je popsán pomocí specifických miRNA např. u ovariálního karcinomu (87).
CAF jsou důležitým producentem růstových faktorů, cytokinů a chemokinů. Tyto molekuly působí jako lokální mediátory v místě svého vzniku, nebo mohou ovlivnit celý organismus. Účastní se řady buněčných dějů, jako je proliferace, diferenciace nebo přežívání buněk. Typicky se vážou na receptory s tyrosinkinázovou aktivitou. Jejich deregulace ovšem může vést k imortalizaci a nekontrolované proliferaci buněk. CAF produkují klasické proonkogenní růstové faktory jako EGF (epidermal growth factor) a HGF (hepatocyte growth factor), proangiogenní faktory jako VEGF (vascular endothelial growth factor), PDGF (platelet-derived growth factor) (34) nebo proteiny BMP-4 (bone morphogenetic protein 4) a IGF-2 (insulin-like growth factor 2), které dediferencují keratinocyty (116. Mimo ně produkují specifické molekuly jako je YAP (yes-associated protein), který podporuje tvorbu metastáz (15), nebo protein WNT16B, který umožnuje nádorovým buňkám uniknout apoptóze a jeho blokace prokazatelně zvyšuje účinnost chemoterapie (122).
V současné době patří protilátky proti růstovým faktorům mezi léčebné modality u pokročilých stadií tumorů. Jako první byla v roce 2004 schválena protilátka anti-VEGF(bevacizumab) u metastáz solidních tumorů kolorektálního karcinomu, nemalobuněčného karcinomu plic, glioblastomu a karcinomu děložního čípku. Mezi další patří například protilátky proti EGFR (cetuximab nebo erlotinib).
CAF jsou také producenty velké skupiny cytokinů. Cytokiny mají pleiotropní účinky na organismus, to znamená, že jeden může působit na více funkcí u různých buněčných typů. Naproti tomu více druhů cytokinů má podobný efekt na jednu buňku. Navzájem jsou schopné působit synergicky nebo antagonisticky, takže plní funkci inhibitorů nebo stimulátorů pro buněčné děje. Z výše uvedených faktů vyplývá, že síť vzájemných interakcí cytokinů je velice složitým a propojeným komplexem (101). Jejich zvýšená produkce může vést ke vzniku autoimunitních onemocnění, ale také přispět k rozvoji nádorového bujení. Mezi nejdůležitější cytokiny, účastnící se nádorové progrese a rezistence k protinádorové terapii, patří chemokiny a interleukiny, podílející se na intenzitě imunitní odpovědi.
Chemokiny atrahují imunitní buňky do oblasti nádorů. Byl prokázán význam CXCL-2 (C-X-C motif ligand 2) při rezistenci ovariálního karcinomu na chemoterapii (paclitaxel a docetaxel) (88). Při použití blokačních protilátek proti tomuto chemokinu je efektivita protinádorové terapie zvýšena (88). Přerušení signální cesty CXCR-4 (C-X-C chemokine receptor type 4) /CXCL-12 zvýšilo senzitivitu chemoterapie (docetaxel) u nádorů prostaty (24), tlustého střeva (52) a plic (14). U CXCL12, CXCL14 (C-X-C motif ligand 14) a CCL5 (C-C motif) ligand 5) je prokázán jejich vliv na vznik metastáz (5, 76). Tyto skutečnosti naznačují, že kromě očekávané protinádorové aktivity mohou buňky imunitního systému podporovat jak růst, tak diseminaci nádoru.
Původně se předpokládalo, že výhradní producenti interleukinů (IL) jsou lymfocyty, ale mohou je produkovat i další buněčné typy, včetně CAF. Hladina IL-1β je zvýšena u pacientů s nádory žaludku a jater (44, 106). Pronádorový účinek IL-6 byl prokázán u nádorů prsu, ovaria, prostaty a plic (12). IL-6 působí také proangiogenně (90) a jeho overexprese je spojena s rezistencí k inhibitoru EGFR (erlotinibu) (41, 145) stejně jako inhibitoru estrogenových receptorů (tamoxifenu) u karcinomu prsu (120). Je popsána korelace mezi zvýšenou hladinou IL-6, IL-8 a progresí tumoru (23, 148). Společná exprese molekul IL-6, IL-8 a CXCL-1 (C-X-C motif ligand 1) je typická pro mikroprostředí SCC a společné působení těchto molekul je zodpovědné za změnu fenotypu zdravých keratinocytů na nízce diferencované nádorové keratinocyty (66). Protilátkami proti IL-6, IL-8 a CXCL-1 jsme schopni této změně zabránit (66).
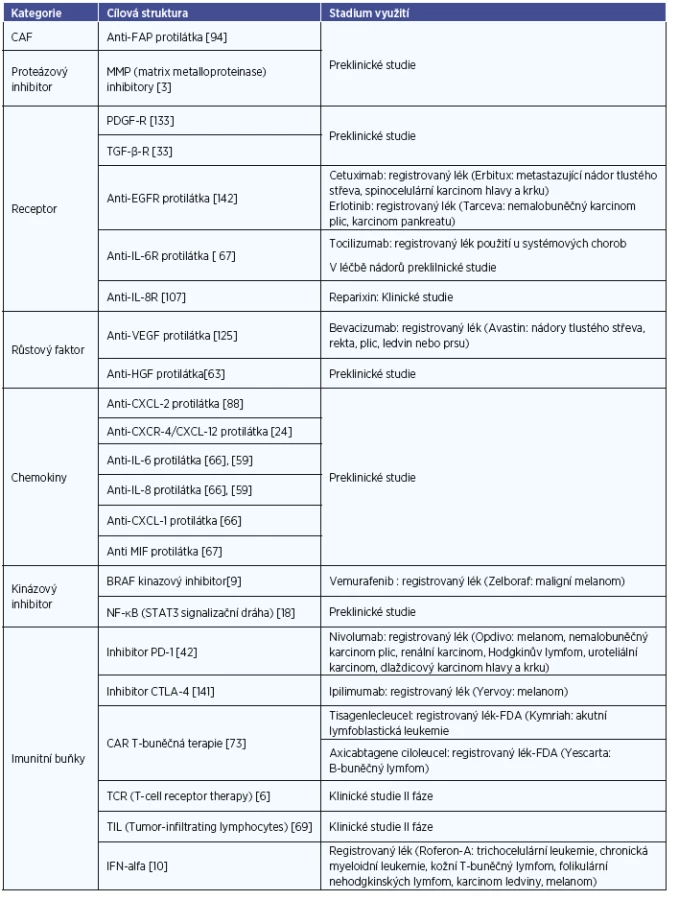
Efekt blokace různých cytokinů byl potvrzen v mnoha preklinických studiích. Můžeme například uvést kombinaci blokace receptorů IL-6 a IL-8 (tocilizumab a reparixin), která u zvířecího modelu signifikantně snížila schopnost tvorby vzdálených metastáz u karcinomu prsu (59). Blokace MIF (macrophage migration inhibitory factor) snížila schopnost angiogeneze a lymfangiogeneze na modelu myšího osteosarkomu (67). Podobně blokací transkripčního faktoru NF-κB (nuclear factor kappa-light-chain) aktivovaného cytokiny a jeho STAT-3 (signal transducer and activator of transcription 3) signalizační dráhy lze zabránit růstu tumoru prostaty (18). Zvláště molekuly IL-6, IL-8, CXCL-1 a TGF-β se zdají být vhodnými kandidáty k dalším experimentům za účelem možné klinické aplikace (8, 65, 117). Nejdůležitější molekuly produkované CAF jsou shrnuty v tabulce 1. Možnost terapeutického ovlivnění je v tabulce 2.

Interakce CAF s nádorovou buňkou se jeví jako nespecifická. Bez ohledu na svůj původ jsou CAF získané z různých typů nádorů schopné u zdravých keratinocytů a buněk karcinomu prsu indukovat expresi keratinů 8, 14 a 19, jejichž společná koexprese je znakem nízce diferencovaných keratinocytů a je špatným prognostickým znakem u pacientů s nádory hlavy a krku a karcinomu prsu (31, 68).
CAF se tak ukazují jako vhodný cíl protinádorové terapie a jejich aktivita je možným faktorem selhání protinádorové léčby cílené pouze na vlastní nádorové buňky (109, 121). Samotné CAF jsou v porovnání s karcinomovými buňkami geneticky stabilnější méně heterogenní skupina s menší šancí na vznik nádorové chemorezistence (20, 61). Jsou však schopné vyvolat mechanismy, kterými nádorové buňky uniknou terapii. Například růstový faktor HGF produkovaný CAF způsobuje rezistenci vůči kinázovým inhibitorům (např.vemurafenib u maligního melanomu) (115, 140).
EXTRACELULÁRNÍ MATRIX A JEJÍ REMODELACE U NÁDORŮ
Aktivitou CAF dochází k neustálé remodelaci ECM, a to jak novotvorbou, tak i její degradací proteázami (11). ECM vytvořená CAF hraje důležitou roli při utváření reaktivního nádorového mikroprostředí (71). ECM je složena z velkého počtu stavebních jednotek jako složitý komplex vzájemně interagujících proteinů a sacharidových molekul. Jednou z komponent ECM nádorů nebo hojící se rány je tenascin-c/w. Stimulační účinek tenascinu-c/w na růst nádorových buněk byl prokázán u mnoha typů nádorů (7). Fibronektin, další z proteinů ECM, se podílí na proliferaci fibroblastů, stimuluje chemotaxi imunokompetentních buněk a podporuje vaskularizaci nádoru (84, 103, 132). V dlaždicovém karcinomu ústní dutiny je přítomnost fibronektinu asociována s přítomností uzlinových metastáz, a tudíž s celkově horší prognózou pro pacienta (84). K molekulám ECM, podporující růst nádoru, patří dále např. lamin (37) a periostin (127) nebo heparan sulfát (45).
GALEKTINY (GAL) JAKO PŘÍKLAD ENDOGENNÍCH LEKTINŮ OVLIVŇUJÍCÍCH NÁDORY
Většina proteinů prochází posttranslační modifikací, glykosilací. Endogenní lektiny jsou bílkoviny v tělech živočichů, které jsou schopné rozlišit specifický cukerný motiv. Cukry jsou tak médiem pro uložení biologické informace, která je čtena (113). Gal (dříve S lektiny) z rodiny endogenních lektinů, se významně podílejí na komunikaci mezi nádorem a nádorovým mikroprostředím. Nemají charakter enzymů či protilátek a jsou schopné specificky rozpoznat β-galaktosidy. Gal se uplatňují v široké škále biologických dějů, kde se podílejí na regulaci proliferace, diferenciace, apoptózy a modulaci mezibuněčné interakce a interakce s ECM. V kancerogenezi je to především gal-1, -3, -7 (74, 99). Přítomnost gal-1 ve stromatu nádorů hlavy a krku je doprovázena zvýšeným výskytem CAF (samy jsou rovněž producenty gal-1) a zároveň je špatným prognostickým faktorem pro vývoj onemocnění (143). Gal-1 je nezávisle na funkci TGF-β schopný transformovat fibroblasty na myofibroblasty a při in vitro kultivaci stimulovat ECM bohatou na fibronectin, tenascin a samotný gal-1 (30, 131). Dále je jeho přítomnost spojena se zvýšenou nádorovou invazivitou, metastazováním buněk a zvýšenou produkcí růstových faktorů, např. VEGF-C (1, 110, 128, 139). Gal-1 umožňuje nádorovým buňkám uniknout protinádorové imunitě pomocí suprese cytotoxických i pomocných T-lymfocytů (16, 130), je také zodpovědný za apoptózu aktivovaných T-lymfocytů (51, 97). Důležitou roli v nádorové progresi hraje i gal-3 (30), který podporuje produkci mnoha prozánětlivých chemokinů jako CCL-2, -5, -8 a -20, nebo CXCL-8 (IL-8) (95) a také modifikuje imunologickou odpověď T-lymfocytů a NK buněk na přítomnost nádorové tkáně (105).
IMUNITNÍ SYSTÉM: BOJOVNÍK PROTI NÁDORU, NEBO JEHO POMOCNÍK
Buňky imunitního systému představují další významnou komponentu nádorového stromatu, jedná se především o makrofágy, lymfocyty a žírné buňky. Imunitní buňky slouží u zdravého organismu k eliminaci buněk s aberantním genomem a zabraňují tak vzniku maligního tumoru (2, 60). Je prokázáno, že nádory (SCC) jsou schopné modifikovat imunologickou odpověď (27, 106). Nádorové buňky mají sníženou expresi hlavního histokompatibilního komplexu MHC-I, tato vlastnost umožňuje uniknout buněčné imunitě (92, 147). Další schopností nádorových buněk je indukce apoptózy T-lymfocytů např. pomocí exprese FasL (Fas ligandu) (138).
Hlavním hráčem protinádorové imunitní odpovědi jsou T - lymfocyty, které spouštějí adaptivní imunitní odpověď a jsou zodpovědně za cytotoxickou destrukci nádorových buněk (138). Můžeme je rozdělit na CD 4+ pozitivní pomocné T-lymfocyty, CD 8+ pozitivní cytotoxické T-lymfocyty a supresorové T-lymfocyty. Zvýšená exprese ligandu PD-L1 (programmed death-ligand), který se váže na PD-1 receptor CD 8+ T-lymfocytů, je prokázána u nádorových buněk a CAF (118). Vlastností PD-L1 je suprese funkce a apoptóza CD 8+ T-lymfocytů [57]. CTLA-4 je dalším důležitým receptorem cytotoxických CD 8+ T-lymfocytů, je zodpovědný za supresi protinádorové imunitní odpovědi (54). Inhibitor PD-1 (nivolumab) je společně s inhibitorem CTLA-4 (ipilimumab) součástí terapie u generalizovaného maligního melanomu s velmi nadějnými výsledky. Cytokiny TGF-β a IL-10 umožňují diferenciaci naivních T-lymfocytů v supresorové T-lymfocyty, které podporují supresi cytotoxických T-lymfocytů (35).
Dendritické buňky jsou nejúčinnější antigen prezentující buňky. Podílejí se na vyzrávání a aktivaci cytotoxických T-lymfocytů a tvoří tak spojovací článek mezi vrozenou a adaptivní imunitní odpovědí. V případě působení TGF-β a IL-10 podporují diferenciaci supresorových T-lymfocytů, a tím zvyšují nádorovou imunotoleranci (50, 55).
Makrofágy přítomné v nádorovém stromatu vznikají z monocytů. Po kontaktu s nádorovými buňkami se diferencují v nádorově asociované makrofágy (Tumor associated macrophages=TAM). Můžeme je rozdělit na 2 typy M1 (prozánětlivé a protinádorové) a M2 (supresorové). M1 přispívají k protinádorové imunitní odpovědí produkcí zánětlivých cytokinů jako IL-12, IL-23 a interferonu-γ (77, 108). Naopak M2 podporují angiogenezi produkcí VEGF, HGF, MMP2 nebo IL-8 (71) a nádorovou imunotoleranci pomocí cytokinů jako TGF-α a IL-10 (32, 70). U SCC je zastoupení M2 spojeno s vyšším stagingem nádorů, metastatickým postižením uzlin s častější extrakaspulární propagací (72, 78). TAM dále produkují MIF (macrophage migration inhibitory factor), který atrahuje neutrofilní granulocytu do místa nádoru, což je spojeno s angiogenezí a vyšší invazivitou nádoru (26, 40).
ZÁVĚR
Nádorové mikroprostředí představuje esenciální složku nádorů. Je tvořeno velmi komplikovanou soustavou rozdílných a vzájemně se ovlivňujících buněčných populací, signálních molekul a ECM. Interakce nádorových buněk s ostatními buňkami ekosystému a jejich produkty je jedním ze zásadních parametrů, který ovlivňuje biologické vlastnosti nádoru. V posledních letech se teoretické znalosti o interakci jednotlivých buněk pomalu přesouvají směrem do klinické praxe. Léky na bázi protilátek proti některým molekulám jsou zařazeny do léčebných schémat pokročilých generalizovaných nádorů s nadějnými výsledky (tab. 2). Tento fakt je velmi perspektivní v dalším zkoumání nádorového mikroprostředí a ve vývoji nových protinádorových léčiv.
Poděkování
Tento přehledný článek vznikl za podpory Karlovy Univerzity v Praze, projekt PROGRES Q28, a Ministerstva zdravotnictví ČR, projekt 15-28933A.
Adresa ke korespondenci:
MUDr. Štěpán Novák
Klinika ORL a chirurgie hlavy a krku 1. LF UK a FN
V Úvalu 84
150 06 Praha 5
e-mail: stepan.novak@fnmotol.cz
Sources
1. Amano, M., Eriksson, H., Manning, J. C. et al.: Tumour suppressor p16(INK4a) - anoikis-favouring decrease in N/O-glycan/cell surface sialylation by down-regulation of enzymes in sialic acid biosynthesis in tandem in a pancreatic carcinoma model. FEBS J, 279, 2012, 21, s. 4062-4080.
2. Amend, S. R. and Pienta, K. J.: Ecology meets cancer biology: the cancer swamp promotes the lethal cancer phenotype. Oncotarget, 6, 2015, 12, s. 9669-78.
3. Appleby, T. C., Greenstein, A. E., Hung, M. et al.: Biochemical characterization and structure determination of a potent, selective antibody inhibitor of human MMP9. J Biol Chem, 292, 2017, 16, s. 6810-6820.
4. Augsten, M., Hagglof, C., Olsson, E. et al.: CXCL14 is an autocrine growth factor for fibroblasts and acts as a multi-modal stimulator of prostate tumor growth. Proc Natl Acad Sci U S A, 106, 2009, 9, s. 3414-9.
5. Augsten, M., Sjoberg, E., Frings, O. et al.: Cancer-associated fibroblasts expressing CXCL14 rely upon NOS1-derived nitric oxide signaling for their tumor-supporting properties. Cancer Res, 74, 2014, 11, s. 2999-3010.
6. Barrett, D. M., Grupp, S. A. and June, C. H.: Chimeric Antigen Receptor - and TCR-Modified T Cells Enter Main Street and Wall Street. J Immunol, 195, 2015, 3, s. 755-61.
7. Berndt, A., Richter, P., Kosmehl, H. and Franz, M.: Tenascin-C and carcinoma cell invasion in oral and urinary bladder cancer. Cell Adh Migr, 9, 2015, 1-2, s. 105-11.
8. Bierie, B., Stover, D. G., Chytil, A. et al.: Transforming growth factor-beta regulates mammary carcinoma cell survival and interaction with the adjacent microenvironment. Cancer Research, 68, 2008, 6, s. 1809-1819.
9. Bollag, G., Tsai, J., Zhang, J. et al.: Vemurafenib: the first drug approved for BRAF-mutant cancer. Nat Rev Drug Discov, 11, 2012, 11, s. 873-86.
10. Bonifazi, F., de Vivo, A., Rosti, G. et al.: Chronic myeloid leukemia and interferon-alpha: a study of complete cytogenetic responders. Blood, 98, 2001, 10, s. 3074-81.
11. Bonnans, C., Chou, J., Werb, Z.: Remodelling the extracellular matrix in development and disease. Nat Rev Mol Cell Biol, 15, 2014, 12, s. 786-801.
12. Bromberg, J. and Wang, T. C.: Inflammation and Cancer: IL-6 and STAT3 Complete the Link. Cancer Cell, 15, 2009, 2, s. 79-80.
13. Buglioni, S., Vici, P., Sergi, D. et al.: Analysis of the hippo transducers TAZ and YAP in cervical cancer and its microenvironment. Oncoimmunology, 5, 2016, 6, s. e1160187.
14. Burger, J. A., Stewart, D. J., Wald, O., Peled, A.: Potential of CXCR4 antagonists for the treatment of metastatic lung cancer. Expert Rev Anticancer Ther, 11, 2011, 4, s. 621-630.
15. Calvo, F., Ege, N., Grande-Garcia, A. et al.: Mechanotransduction and YAP-dependent matrix remodelling is required for the generation and maintenance of cancer-associated fibroblasts. Nat Cell Biol, 15, 2013, 6, s. 637-646.
16. Cedeno-Laurent, F., Dimitroff, C. J.: Galectins and their ligands: negative regulators of anti-tumor immunity. Glycoconj J, 29, 2012, 8-9, s. 619-625.
17. Cirri, P. and Chiarugi, P.: Cancer associated fibroblasts: the dark side of the coin. Am J Cancer Res, 1, 2011, 4, s. 482-497.
18. Civenni, G., Longoni, N., Costales, P. et al.: EC-70124, a Novel Glycosylated Indolocarbazole Multikinase Inhibitor, Reverts Tumorigenic and Stem Cell Properties in Prostate Cancer by Inhibiting STAT3 and NF-kappaB. Mol Cancer Ther, 15, 2016, 5, s. 806-818.
19. Clayton, A., Evans, R. A., Pettit, E. et al.: Cellular activation through the ligation of intercellular adhesion molecule-1. J Cell Sci, 111 ( Pt 4), 1998, s. 443-453.
20. Correia, A. L., Bissell, M. J.: The tumor microenvironment is a dominant force in multidrug resistance. Drug Resist Updat, 15, 2012, 1-2, s. 39-49.
21. Curry, J. M., Sprandio, J., Cognetti, D. et al.: Tumor microenvironment in head and neck squamous cell carcinoma. Semin Oncol, 41, 2014, 2, s. 217-234.
22. De Kruijf, E. M., van Nes, J. G., van de Velde, C. J. et al.: Tumor-stroma ratio in the primary tumor is a prognostic factor in early breast cancer patients, especially in triple-negative carcinoma patients. Breast Cancer Res Treat, 125, 2011, 3, s. 687-696.
23. Dhawan, P., Richmond, A.: Role of CXCL1 in tumorigenesis of melanoma. J Leukoc Biol, 72, 2002, 1, s. 9-18.
24. Domanska, U. M., Timmer-Bosscha, H., Nagengast, W. B. et al.: CXCR4 inhibition with AMD3100 sensitizes prostate cancer to docetaxel chemotherapy. Neoplasia, 14, 2012, 8, s. 709-718.
25. Du, Y. E., Tu, G., Yang, G. et al.: MiR-205/YAP1 in Activated Fibroblasts of Breast Tumor Promotes VEGF-independent Angiogenesis through STAT3 Signaling. Theranostics, 7, 2017, 16, s. 3972-3988.
26. Dumitru, C. A., Gholaman, H., Trellakis, S. et al.: Tumor-derived macrophage migration inhibitory factor modulates the biology of head and neck cancer cells via neutrophil activation. Int J Cancer, 129, 2011, 4, s. 859-869.
27. Duray, A., Demoulin, S., Hubert, P., Delvenne, P., Saussez, S.: Immune suppression in head and neck cancers: a review. Clin Dev Immunol, 2010, 2010, s. 701657.
28. Dvorak, H. F.: Tumors: wounds that do not heal. Similarities between tumor stroma generation and wound healing. N Engl J Med, 315, 1986, 26, s. 1650-1659.
29. Dvorankova, B., Smetana, K., Jr., Rihova, B. et al.: Cancer-associated fibroblasts are not formed from cancer cells by epithelial-to-mesenchymal transition in nu/nu mice. Histochem Cell Biol, 143, 2015, 5, s. 463-469.
30. Dvorankova, B., Szabo, P., Lacina, L. et al.: Human galectins induce conversion of dermal fibroblasts into myofibroblasts and production of extracellular matrix: potential application in tissue engineering and wound repair. Cells Tissues Organs, 194, 2011, 6, s. 469-480.
31. Dvorankova, B., Szabo, P., Lacina, L. et al.: Fibroblasts prepared from different types of malignant tumors stimulate expression of luminal marker keratin 8 in the EM-G3 breast cancer cell line. Histochem Cell Biol, 137, 2012, 5, s. 679-685.
32. El-Rouby, D. H.: Association of macrophages with angiogenesis in oral verrucous and squamous cell carcinomas. J Oral Pathol Med, 39, 2010, 7, s. 559-564.
33. Faivre, S. J., Santoro, A., Gane, E. et al.: A phase 2 study of galunisertib, a novel transforming growth factor-beta (TGF-beta) receptor I kinase inhibitor in patients with advanced hepatocellular carcinoma (HCC) and low serum alpha fetoprotein (AFP). Journal of Clinical Oncology, 34, 2016, 15.
34. Ferrara, N.: Pathways mediating VEGF-independent tumor angiogenesis. Cytokine Growth Factor Rev, 21, 2010, 1, s. 21-26.
35. Ferris, R. L., Whiteside, T. L. and Ferrone, S.: Immune escape associated with functional defects in antigen-processing machinery in head and neck cancer. Clin Cancer Res, 12, 2006, 13, s. 3890-3895.
36. Fidler, I. J.: The pathogenesis of cancer metastasis: the ‚seed and soil‘ hypothesis revisited. Nat Rev Cancer, 3, 2003, 6, s. 453-458.
37. Franz, M., Hansen, T., Richter, P. et al.: Complex formation of the laminin-5 gamma2 chain and large unspliced tenascin-C in oral squamous cell carcinoma in vitro and in situ: implications for sequential modulation of extracellular matrix in the invasive tumor front. Histochem Cell Biol, 126, 2006, 1, s. 125-131.
38. Fujita, H., Ohuchida, K., Mizumoto, K. et al.: Tumor-stromal interactions with direct cell contacts enhance proliferation of human pancreatic carcinoma cells. Cancer Sci, 100, 2009, 12, s. 2309-2317.
39. Fukui, H., Zhang, X., Sun, C. et al.: IL-22 produced by cancer-associated fibroblasts promotes gastric cancer cell invasion via STAT3 and ERK signaling. Br J Cancer, 111, 2014, 4, s. 763-771.
40. Galdiero, M. R., Garlanda, C., Jaillon, S., Marone, G., Mantovani, A.: Tumor associated macrophages and neutrophils in tumor progression. J Cell Physiol, 228, 2013, 7, s. 1404-1412.
41. Gao, S. P., Mark, K. G., Leslie, K. et al.: Mutations in the EGFR kinase domain mediate STAT3 activation via IL-6 production in human lung adenocarcinomas. J Clin Invest, 117, 2007, 12, s. 3846-3856.
42. Gettinger, S., Horn, L., Jackman, D. et al.: Five-year follow-up of nivolumab in previously treated advanced non-small-cell lung cancer: Results from the CA209-003 study. J Clin Oncol, 36, 2018, 17, s. 1675-1684.
43. Gialeli, C., Nikitovic, D., Kletsas, D. et al.: PDGF/PDGFR signaling and targeting in cancer growth and progression: Focus on tumor microenvironment and cancer-associated fibroblasts. Curr Pharm Des, 20, 2014, 17, s. 2843-2848.
44. Glas, J., Toeroek, H. P., Schneider, A. et al.: Allele 2 of the interleukin-1 receptor antagonist gene: An independent genetic risk factor in early gastric cancer. Gastroenterology, 126, 2004, 4, s. A453-A453.
45. Gomes, A. M., Stelling, M. P., Pavao, M. S.: Heparan sulfate and heparanase as modulators of breast cancer progression. Biomed Res Int, 2013, 2013, s. 852093.
46. Gore, J., Korc, M.: Pancreatic cancer stroma: friend or foe? Cancer Cell, 25, 2014, 6, s. 711-712.
47. Gudleviciene, Z., Didziapetriene, J., Mackeviciene, I. et al.: Prevalence of human papillomaviruses in patients with head and neck squamous cell carcinoma in Lithuania and Belarus. J Med Virol, 86, 2014, 3, s. 531-535.
48. Hamburger, A. W., Salmon, S. E.: Primary bioassay of human tumor stem cells. Science, 197, 1977, 4302, s. 461-463.
49. Hammarstedt, L., Lindquist, D., Dahlstrand, H. et al.: Human papillomavirus as a risk factor for the increase in incidence of tonsillar cancer. Int J Cancer, 119, 2006, 11, s. 2620-2623.
50. Hawiger, D., Inaba, K., Dorsett, Y. et al.: Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J Exp Med, 194, 2001, 6, s. 769-779.
51. He, J., Baum, L. G.: Presentation of galectin-1 by extracellular matrix triggers T cell death. J Biol Chem, 279, 2004, 6, s. 4705-4712.
52. Heckmann, D., Maier, P., Laufs, S. et al.: CXCR4 Expression and treatment with SDF-1alpha or plerixafor nodulate proliferation and chemosensitivity of colon cancer cells. Transl Oncol, 6, 2013, 2, s. 124-232.
53. Honjo, Y., Inohara, H., Akahani, S. et al.: Expression of cytoplasmic galectin-3 as a prognostic marker in tongue carcinoma. Clin Cancer Res, 6, 2000, 12, s. 4635-4640.
54. Hoos, A., Ibrahim, R., Korman, A. et al.: Development of ipilimumab: contribution to a new paradigm for cancer immunotherapy. Semin Oncol, 37, 2010, 5, s. 533-46.
55. Charalambous, A., Oks, M., Nchinda, G., Yamazaki, S. and Steinman, R. M.: Dendritic cell targeting of survivin protein in a xenogeneic form elicits strong CD4+ T cell immunity to mouse survivin. J Immunol, 177, 2006, 12, s. 8410-8421.
56. Chatzistamou, I., Dioufa, N., Trimis, G. et al.: p21/waf1 and smooth-muscle actin alpha expression in stromal fibroblasts of oral cancers. Cell Oncol (Dordr), 34, 2011, 5, s. 483-488.
57. Cho, Y. A., Yoon, H. J., Lee, J. I., Hong, S. P., Hong, S. D.: Relationship between the expressions of PD-L1 and tumor-infiltrating lymphocytes in oral squamous cell carcinoma. Oral Oncol, 47, 2011, 12, s. 1148-1153.
58. Jansson, S., Aaltonen, K., Bendahl, P. O. et al.: The PDGF pathway in breast cancer is linked to tumour aggressiveness, triple-negative subtype and early recurrence. Breast Cancer Res Treat, 169, 2018, 2, s. 231-241.
59. Jayatilaka, H., Tyle, P., Chen, J. J. et al.: Synergistic IL-6 and IL-8 paracrine signalling pathway infers a strategy to inhibit tumour cell migration. Nat Commun, 8, 2017, s. 15584.
60. Kareva, I.: What can ecology teach us about cancer? Transl Oncol, 4, 2011, 5, s. 266-270.
61. Kerbel, R. S.: A cancer therapy resistant to resistance. Nature, 390, 1997, 6658, s. 335-336.
62. Kilvaer, T. K., Rakaee, M., Hellevik, T. et al.: Tissue analyses reveal a potential immune-adjuvant function of FAP-1 positive fibroblasts in non-small cell lung cancer. PLoS One, 13, 2018, 2, s. e0192157.
63. Kim, H., Hong, S. H., Kim, J. Y. et al.: Preclinical development of a humanized neutralizing antibody targeting HGF. Exp Mol Med, 49, 2017, 3, s. e309.
64. Kim, H. M., Jung, W. H. and Koo, J. S.: Expression of cancer-associated fibroblast related proteins in metastatic breast cancer: an immunohistochemical analysis. J Transl Med, 13, 2015, s. 222.
65. Kolar, M., Szabo, P., Dvorankova, B. et al.: Upregulation of IL-6, IL-8 and CXCL-1 production in dermal fibroblasts by normal/malignant epithelial cells in vitro: Immunohistochemical and transcriptomic analyses. Biology of the Cell, 104, 2012, 12, s. 738-751.
66. Kolar, M., Szabo, P., Dvorankova, B. et al.: Upregulation of IL-6, IL-8 and CXCL-1 production in dermal fibroblasts by normal/malignant epithelial cells in vitro: Immunohistochemical and transcriptomic analyses. Biol Cell, 104, 2012, 12, s. 738-751.
67. Kubota, Y., Takubo, K., Shimizu, T. et al.: M-CSF inhibition selectively targets pathological angiogenesis and lymphangiogenesis. J Exp Med, 206, 2009, 5, s. 1089-1102.
68. Lacina, L., Dvorankova, B., Smetana, K., Jr. et al.: Marker profiling of normal keratinocytes identifies the stroma from squamous cell carcinoma of the oral cavity as a modulatory microenvironment in co-culture. Int J Radiat Biol, 83, 2007, 11-12, s. 837-848.
69. Lee, H. J., Kim, Y. A., Sim, C. K. et al.: Expansion of tumor-infiltrating lymphocytes and their potential for application as adoptive cell transfer therapy in human breast cancer. Oncotarget, 8, 2017, 69, s. 113345-113359.
70. Li, C., Shintani, S., Terakado, N., Nakashiro, K., Hamakawa, H.: Infiltration of tumor-associated macrophages in human oral squamous cell carcinoma. Oncol Rep, 9, 2002, 6, s. 1219-1223.
71. Li, H., Fan, X., Houghton, J.: Tumor microenvironment: the role of the tumor stroma in cancer. J Cell Biochem, 101, 2007, 4, s. 805-815.
72. Liu, S. Y., Chang, L. C., Pan, L. F. et al.: Clinicopathologic significance of tumor cell-lined vessel and microenvironment in oral squamous cell carcinoma. Oral Oncol, 44, 2008, 3, s. 277-285.
73. Liu, Y., Chen, X., Han, W., Zhang, Y.: Tisagenlecleucel, an approved anti-CD19 chimeric antigen receptor T-cell therapy for the treatment of leukemia. Drugs Today (Barc), 53, 2017, 11, s. 597-608.
74. Lohr, M., Lensch, M., Andre, S. et al.: Murine homodimeric adhesion/growth-regulatory galectins-1, -2 and -7: comparative profiling of gene/ promoter sequences by database mining, of expression by RT-PCR/immunohistochemistry and of contact sites for carbohydrate ligands by computational chemistry. Folia Biol (Praha), 53, 2007, 4, s. 109-128.
75. Lorusso, G., Ruegg, C.: The tumor microenvironment and its contribution to tumor evolution toward metastasis. Histochem Cell Biol, 130, 2008, 6, s. 1091-1103.
76. Luker, K. E., Lewin, S. A., Mihalko, L. A. et al.: Scavenging of CXCL12 by CXCR7 promotes tumor growth and metastasis of CXCR4-positive breast cancer cells. Oncogene, 31, 2012, 45, s. 4750-4758.
77. Mantovani, A., Bottazzi, B., Colotta, F., Sozzani, S., Ruco, L.: The origin and function of tumor-associated macrophages. Immunol Today, 13, 1992, 7, s. 265-270.
78. Marcus, B., Arenberg, D., Lee, J. et al.: Prognostic factors in oral cavity and oropharyngeal squamous cell carcinoma. Cancer, 101, 2004, 12, s. 2779-2787.
79. Marsh, T., Pietras, K., McAllister, S. S.: Fibroblasts as architects of cancer pathogenesis. Biochim Biophys Acta, 1832, 2013, 7, s. 1070-1078.
80. Martin, F. T., Dwyer, R. M., Kelly, J. et al.: Potential role of mesenchymal stem cells (MSCs) in the breast tumour microenvironment: stimulation of epithelial to mesenchymal transition (EMT). Breast Cancer Res Treat, 124, 2010, 2, s. 317-326.
81. Melaku, Y. A., Appleton, S. L., Gill, T. K. et al.: Incidence, prevalence, mortality, disability-adjusted life years and risk factors of cancer in Australia and comparison with OECD countries, 1990-2015: findings from the Global Burden of Disease Study 2015. Cancer Epidemiol, 52, 2018, s. 43-54.
82. Mesker, W. E., Junggeburt, J. M., Szuhai, K. et al.: The carcinoma-stromal ratio of colon carcinoma is an independent factor for survival compared to lymph node status and tumor stage. Cell Oncol, 29, 2007, 5, s. 387-398.
83. Metwaly, H., Maruyama, S., Yamazaki, M. et al.: Parenchymal-stromal switching for extracellular matrix production on invasion of oral squamous cell carcinoma. Hum Pathol, 43, 2012, 11, s. 1973-1981.
84. Mhawech, P., Dulguerov, P., Assaly, M., Ares, C., Allal, A. S.: EB-D fibronectin expression in squamous cell carcinoma of the head and neck. Oral Oncol, 41, 2005, 1, s. 82-88.
85. Micke, P., Kappert, K., Ohshima, M. et al.: In situ identification of genes regulated specifically in fibroblasts of human basal cell carcinoma. J Invest Dermatol, 127, 2007, 6, s. 1516-1523.
86. Mishra, P. J., Mishra, P. J., Glod, J. W., Banerjee, D.: Mesenchymal stem cells: flip side of the coin. Cancer Res, 69, 2009, 4, s. 1255-1258.
87. Mitra, A. K., Zillhardt, M., Hua, Y. et al.: MicroRNAs reprogram normal fibroblasts into cancer-associated fibroblasts in ovarian cancer. Cancer Discov, 2, 2012, 12, s. 1100-1118.
88. Moisan, F., Francisco, E. B., Brozovic, A. et al.: Enhancement of paclitaxel and carboplatin therapies by CCL2 blockade in ovarian cancers. Mol Oncol, 8, 2014, 7, s. 1231-1239.
89. Motlik, J., Klima, J., Dvorankova, B., Smetana, K., Jr.: Porcine epidermal stem cells as a biomedical model for wound healing and normal/malignant epithelial cell propagation. Theriogenology, 67, 2007, 1, s. 105-111.
90. Nagasaki, T., Hara, M., Nakanishi, H. et al.: Interleukin-6 released by colon cancer-associated fibroblasts is critical for tumour angiogenesis: anti-interleukin-6 receptor antibody suppressed angiogenesis and inhibited tumour-stroma interaction. Br J Cancer, 110, 2014, 2, s. 469-478.
91. Nasman, A., Attner, P., Hammarstedt, L. et al.: Incidence of human papillomavirus (HPV) positive tonsillar carcinoma in Stockholm, Sweden: an epidemic of viral-induced carcinoma? Int J Cancer, 125, 2009, 2, s. 362-366.
92. Ogino, T., Shigyo, H., Ishii, H. et al.: HLA class I antigen down-regulation in primary laryngeal squamous cell carcinoma lesions as a poor prognostic marker. Cancer Res, 66, 2006, 18, s. 9281-9289.
93. Orimo, A., Gupta, P. B., Sgroi, D. C. et al.: Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell, 121, 2005, 3, s. 335-348.
94. Ostermann, E., Garin-Chesa, P., Heider, K. H. et al.: Effective immunoconjugate therapy in cancer models targeting a serine protease of tumor fibroblasts. Clin Cancer Res, 14, 2008, 14, s. 4584-4592.
95. Papaspyridonos, M., McNeill, E., de Bono, J. P. et al.: Galectin-3 is an amplifier of inflammation in atherosclerotic plaque progression through macrophage activation and monocyte chemoattraction. Arterioscler Thromb Vasc Biol, 28, 2008, 3, s. 433-440.
96. Perez-Losada, J. and Balmain, A.: Stem-cell hierarchy in skin cancer. Nat Rev Cancer, 3, 2003, 6, s. 434-443.
97. Perillo, N. L., Pace, K. E., Seilhamer, J. J., Baum, L. G.: Apoptosis of T cells mediated by galectin-1. Nature, 378, 1995, 6558, s. 736-739.
98. Plzak, J., Lacina, L., Chovanec, M. et al.: Epithelial-stromal interaction in squamous cell epithelium-derived tumors: an important new player in the control of tumor biological properties. Anticancer Res, 30, 2010, 2, s. 455-462.
99. Plzak, J., Smetana, K., Jr., Hrdlickova, E. et al.: Expression of galectin-3-reactive ligands in squamous cancer and normal epithelial cells as a marker of differentiation. Int J Oncol, 19, 2001, 1, s. 59-64.
100. Polyak, K., Haviv, I., Campbell, I. G.: Co-evolution of tumor cells and their microenvironment. Trends Genet, 25, 2009, 1, s. 30-8.
101. Rakesh, K., Agrawal, D. K.: Controlling cytokine signaling by constitutive inhibitors. Biochemical Pharmacology, 70, 2005, 5, s. 649-657.
102. Rasanen, K., Vaheri, A.: Activation of fibroblasts in cancer stroma. Exp Cell Res, 316, 2010, 17, s. 2713-22.
103. Ritzenthaler, J. D., Han, S. and Roman, J.: Stimulation of lung carcinoma cell growth by fibronectin-integrin signalling. Mol Biosyst, 4, 2008, 12, s. 1160-9.
104. Rupp, C., Scherzer, M., Rudisch, A. et al.: IGFBP7, a novel tumor stroma marker, with growth-promoting effects in colon cancer through a paracrine tumor-stroma interaction. Oncogene, 34, 2015, 7, s. 815-25.
105. Ruvolo, P. P.: Galectin 3 as a guardian of the tumor microenvironment. Biochim Biophys Acta, 1863, 2016, 3, s. 427-437.
106. Sakurai, T., He, G., Matsuzawa, A. et al.: Hepatocyte necrosis induced by oxidative stress and IL-1 alpha release mediate carcinogen-induced compensatory proliferation and liver tumorigenesis. Cancer Cell, 14, 2008, 2, s. 156-165.
107. Schott, A. F., Goldstein, L. J., Cristofanilli, M. et al.: Phase Ib Pilot Study to Evaluate Reparixin in Combination with Weekly Paclitaxel in Patients with HER-2-Negative Metastatic Breast Cancer. Clin Cancer Res, 23, 2017, 18, s. 5358-5365.
108. Sica, A., Schioppa, T., Mantovani, A., Allavena, P.: Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: potential targets of anti-cancer therapy. Eur J Cancer, 42, 2006, 6, s. 717-27.
109. Slany, A., Bileck, A., Muqaku, B., Gerner, C.: Targeting breast cancer-associated fibroblasts to improve anti-cancer therapy. Breast, 24, 2015, 5, s. 532-8.
110. Smetana, K., Jr., Andre, S., Kaltner, H., Kopitz, J., Gabius, H. J.: Context-dependent multifunctionality of galectin-1: a challenge for defining the lectin as therapeutic target. Expert Opin Ther Targets, 17, 2013, 4, s. 379-92.
111. Smetana, K., Jr., Lacina, L., Szabo, P. et al.: Ageing as an Important Risk Factor for Cancer. Anticancer Res, 36, 2016, 10, s. 5009-5017.
112. Smith, E. M., Ritchie, J. M., Summersgill, K. F. et al.: Age, sexual behavior and human papillomavirus infection in oral cavity and oropharyngeal cancers. Int J Cancer, 108, 2004, 5, s. 766-72.
113. Solis, D., Bovin, N. V., Davis, A. P. et al.: A guide into glycosciences: How chemistry, biochemistry and biology cooperate to crack the sugar code. Biochim Biophys Acta, 1850, 2015, 1, s. 186-235.
114. Song, I. Y., Balmain, A.: Cellular reprogramming in skin cancer. Semin Cancer Biol, 32, 2015, s. 32-9.
115. Straussman, R., Morikawa, T., Shee, K. et al.: Tumour micro-environment elicits innate resistance to RAF inhibitors through HGF secretion. Nature, 487, 2012, 7408, s. 500-4.
116. Strnad, H., Lacina, L., Kolar, M. et al.: Head and neck squamous cancer stromal fibroblasts produce growth factors influencing phenotype of normal human keratinocytes. Histochemistry and Cell Biology, 133, 2010, 2, s. 201-211.
117. Strnad, H., Lacina, L., Kolar, M. et al.: Head and neck squamous cancer stromal fibroblasts produce growth factors influencing phenotype of normal human keratinocytes. Histochem Cell Biol, 133, 2010, 2, s. 201-11.
118. Strome, S. E., Dong, H., Tamura, H. et al.: B7-H1 blockade augments adoptive T-cell immunotherapy for squamous cell carcinoma. Cancer Res, 63, 2003, 19, s. 6501-5.
119. Sulic, S., Panic, L., Dikic, I., Volarevic, S.: Deregulation of cell growth and malignant transformation. Croat Med J, 46, 2005, 4, s. 622-38.
120. Sun, X., Mao, Y., Wang, J. et al.: IL-6 secreted by cancer-associated fibroblasts induces tamoxifen resistance in luminal breast cancer. Oncogene, 2014, s.
121. Sun, Y.: Translational horizons in the tumor microenvironment: harnessing breakthroughs and targeting cures. Med Res Rev, 35, 2015, 2, s. 408-436.
122. Sun, Y., Campisi, J., Higano, C. et al.: Treatment-induced damage to the tumor microenvironment promotes prostate cancer therapy resistance through WNT16B. Nat Med, 18, 2012, 9, s. 1359-1368.
123. Szabo, P., Kolar, M., Dvorankova, B. et al.: Mouse 3T3 fibroblasts under the influence of fibroblasts isolated from stroma of human basal cell carcinoma acquire properties of multipotent stem cells. Biol Cell, 103, 2011, 5, s. 233-48.
124. Szabo, P., Valach, J., Smetana, K., Jr., Dvorankova, B.: Comparative analysis of IL-8 and CXCL-1 production by normal and cancer stromal fibroblasts. Folia Biol (Praha), 59, 2013, 3, s. 134-7.
125. Tewari, K. S., Sill, M. W., Penson, R. T. et al.: Bevacizumab for advanced cervical cancer: final overall survival and adverse event analysis of a randomised, controlled, open-label, phase 3 trial (Gynecologic Oncology Group 240). Lancet, 390, 2017, 10103, s. 1654-1663.
126. Thiery, J. P., Acloque, H., Huang, R. Y. and Nieto, M. A.: Epithelial-mesenchymal transitions in development and disease. Cell, 139, 2009, 5, s. 871-90.
127. Tilman, G., Mattiussi, M., Brasseur, F., van Baren, N. and Decottignies, A.: Human periostin gene expression in normal tissues, tumors and melanoma: evidences for periostin production by both stromal and melanoma cells. Mol Cancer, 6, 2007, s. 80.
128. Timoshenko, A. V., Kaltner, H., Andre, S., Gabius, H. J., Lala, P. K.: Differential stimulation of VEGF-C production by adhesion/growth-regulatory galectins and plant lectins in human breast cancer cells. Anticancer Res, 30, 2010, 12, s. 4829-33.
129. Tommelein, J., Verset, L., Boterberg, T. et al.: Cancer-associated fibroblasts connect metastasis-promoting communication in colorectal cancer. Front Oncol, 5, 2015, s. 63.
130. Toscano, M. A., Bianco, G. A., Ilarregui, J. M. et al.: Differential glycosylation of TH1, TH2 and TH-17 effector cells selectively regulates susceptibility to cell death. Nat Immunol, 8, 2007, 8, s. 825-34.
131. Valach, J., Fik, Z., Strnad, H. et al.: Smooth muscle actin-expressing stromal fibroblasts in head and neck squamous cell carcinoma: increased expression of galectin-1 and induction of poor prognosis factors. Int J Cancer, 131, 2012, 11, s. 2499-508.
132. Van Obberghen-Schilling, E., Tucker, R. P., Saupe, F. et al.: Fibronectin and tenascin-C: accomplices in vascular morphogenesis during development and tumor growth. Int J Dev Biol, 55, 2011, 4-5, s. 511-25.
133. Wagner, A. J., Kindler, H., Gelderblom, H. et al.: A phase II study of a human anti-PDGFRalpha monoclonal antibody (olaratumab, IMC-3G3) in previously treated patients with metastatic gastrointestinal stromal tumors. Ann Oncol, 28, 2017, 3, s. 541-546.
134. Walker, M. R., Patel, K. K. and Stappenbeck, T. S.: The stem cell niche. J Pathol, 217, 2009, 2, s. 169-80.
135. Wang, K., Ma, W., Wang, J. et al.: Tumor-stroma ratio is an independent predictor for survival in esophageal squamous cell carcinoma. J Thorac Oncol, 7, 2012, 9, s. 1457-61.
136. Wang, Z., Liu, H., Zhao, R. et al.: Tumor-stroma ratio is an independent prognostic factor of non-small cell lung cancer. Zhongguo Fei Ai Za Zhi, 16, 2013, 4, s. 191-6.
137. Warnakulasuriya, S.: Global epidemiology of oral and oropharyngeal cancer. Oral Oncol, 45, 2009, 4-5, s. 309-16.
138. Whiteside, T. L.: Immunobiology of head and neck cancer. Cancer Metastasis Rev, 24, 2005, 1, s. 95-105.
139. Wiest, I., Alexiou, C., Kuhn, C. et al.: Expression of different carbohydrate tumour markers and galectins 1 and 3 in normal squamous and malignant epithelia of the upper aaerodigestive tract. Anticancer Res, 32, 2012, 5, s. 2023-9.
140. Wilson, T. R., Fridlyand, J., Yan, Y. et al.: Widespread potential for growth-factor-driven resistance to anticancer kinase inhibitors. Nature, 487, 2012, 7408, s. 505-9.
141. Wolchok, J. D., Rollin, L. and Larkin, J.: Nivolumab and Ipilimumab in Advanced Melanoma. N Engl J Med, 377, 2017, 25, s. 2503-2504.
142. Wong, S. F.: Cetuximab: an epidermal growth factor receptor monoclonal antibody for the treatment of colorectal cancer. Clin Ther, 27, 2005, 6, s. 684-94.
143. Wu, M. H., Hong, H. C., Hong, T. M. et al.: Targeting galectin-1 in carcinoma-associated fibroblasts inhibits oral squamous cell carcinoma metastasis by downregulating MCP-1/CCL2 expression. Clin Cancer Res, 17, 2011, 6, s. 1306-16.
144. Wu, X., Ruan, L., Yang, Y. and Mei, Q.: Analysis of gene expression changes associated with human carcinoma-associated fibroblasts in non-small cell lung carcinoma. Biol Res, 50, 2017, 1, s. 6.
145. Yao, Z., Fenoglio, S., Gao, D. C. et al.: TGF-beta IL-6 axis mediates selective and adaptive mechanisms of resistance to molecular targeted therapy in lung cancer. Proc Natl. Acad. Sci U S A, 107, 2010, 35, s. 15535-40.
146. Yeh, C. R., Slavin, S., Da, J. et al.: Estrogen receptor alpha in cancer associated fibroblasts suppresses prostate cancer invasion via reducing CCL5, IL6 and macrophage infiltration in the tumor microenvironment. Mol Cancer, 15, 2016, s. 7.
147. Young, M. R.: Protective mechanisms of head and neck squamous cell carcinomas from immune assault. Head Neck, 28, 2006, 5, s. 462-70.
148. Yurkovetsky, Z. R., Kirkwood, J. M., Edington, H. D. et al.: Multiplex analysis of serum cytokines in melanoma patients treated with interferon-alpha2b. Clin. Cancer Res., 13, 2007, 8, s. 2422-8.
149. Zhang, X. H., Jin, X., Malladi, S. et al.: Selection of bone metastasis seeds by mesenchymal signals in the primary tumor stroma. Cell, 154, 2013, 5, s. 1060-1073.
Labels
Audiology Paediatric ENT ENT (Otorhinolaryngology)Article was published in
Otorhinolaryngology and Phoniatrics

2019 Issue 1
Most read in this issue
- Congenital Stridor
- Asymmetric Hearing Loss
- Cholesteatoma Behind an Intact Tympanic Membrane – Retrospective Study
- Tumor Microenvironment