
Vliv aktivního pohybu a pasivního sledování stejného pohybu na elektrickou mozkovou aktivitu
The Effect of Active Movement and Passive Monitoring of the Same Movement on Electric Brain Activity
Introduction:
Mirror neurons are specific association neurons that are active both during a certain movement and during the observation of such movement. It can be assumed that mirror neurons play an important role in the ability to imitate and are an important condition for the rehabilitation of the locomotor system.
Methods:
The aim of our work was to find out whether and how the electrical brain activity is evaluated using the sLORETA display between the actual active movement of a particular motion and the passive tracking of the same motion performed by the therapist or the same motion in the 2D video monitored in the virtual environment.
Results:
The study was attended by 12 healthy, college students, 8 women and 4 men, whose average age was 24 years. Electroencephalographic activity was registered using the Natus Neurology Nicolet EEG Wireless 32/64 Amplifier, a 32-channel telemetric EEG device. (USA). Statistically significant differences in the significance level of p≤0.05 were found in the alpha-1, 2 and beta-2 band when comparing active movement (ACTIVE) versus resting EEG (PRE OE), passive motion monitoring by the therapist PASSIVE versus resting EEG activity with open eyes (PRE OE) and passive 2D motion video viewing (VIDEO) versus resting EEG open-eye activity (PRE OE).
Conclusion:
The results show that there is a great similarity in brain activity in monitoring the movement of the therapist and in the actual implementation of the same movement by the patient. This finding completely corresponds to the existence of mirror neurons and the results of published studies. The second finding is the finding of a difference in brain activity between real-world motion tracking and tracking the same motion in 2D video in virtual reality glasses.
KEYWORDS
motoric, sLORETA, mirror neurons, virtual reality
Authors:
D. Pánek 1; T. Nováková 1; M. Brunovský 2; J. Košťálová 1; D. Pavlů 1
Authors‘ workplace:
Katedra fyzioterapie, Fakulta tělesné výchovy a sportu, Univerzita Karlova, Praha
1; Národní ústav duševního zdraví, Klecany
2
Published in:
Rehabil. fyz. Lék., 25, 2018, No. 4, pp. 152-157.
Category:
Original Papers
Overview
Úvod:
Zrcadlové neurony jsou specifické asociační neurony, které jsou aktivní jak během provádění určitého pohybu, tak i během pozorování stejného pohybu. Lze předpokládat, že zrcadlové neurony hrají důležitou roli ve schopnosti imitace a jsou důležitou podmínkou rehabilitace pohybového systému.
Metody:
Cílem naší práce bylo zjistit, zda a jak se liší elektrická mozková aktivita hodnocená pomocí sLORETA zobrazení mezi vlastním aktivním provedením určitého pohybu a pasivním sledováním stejného pohybu, který provádí před probandem terapeut, nebo sledování stejného pohybu ve 2D videu sledovaného ve virtuálním prostředí.
Výsledky:
Studie se zúčastnilo 12 zdravých, vysokoškolských studentů, 8 žen a 4 muži, jejichž průměrný věk byl 24 let. Elektroencefalografická aktivita byla registrována pomocí telemetrického 32kanálového EEG přístroje Nicolet EEG wireless 32/64 Amplifier firmy Natus Neurology, Inc. (USA). Statisticky signifikantní diference na hladině významnosti p≤0,05 byly zjištěny v pásmu alfa-1, 2 a beta-2 při porovnání aktivního provedení pohybu (ACTIVE) versus klidová EEG aktivita s otevřenýma očima (PRE OE), pasivního sledování pohybu prováděného terapeutem (PASSIVE) versus klidová EEG aktivita s otevřenýma očima (PRE OE) a pasivního sledování 2D videa s prováděním pohybu terapeutem (VIDEO) versus klidová EEG aktivita s otevřenýma očima (PRE OE).
Závěr:
Výsledky ukazují, že existuje velká podobnost mozkové aktivity při sledování pohybu terapeuta a při vlastním provádění stejného pohybu pacientem. Tento nález zcela koresponduje s existencí zrcadlových neuronů a s výsledky publikovaných studií. Druhým zjištěním je nález diference v mozkové aktivitě mezi sledováním pohybu u reálného terapeuta a sledováním stejného pohybu v 2D videu v brýlích pro virtuální realitu.
klíčová slova
motorika, sLORETA, zrcadlové neurony, virtuální realita
ÚVOD
Ve fyzioterapeutické praxi je zcela běžné, že terapeut předvádí sám pohyb a pacient ho za současné korekce terapeuta zopakuje. Efekt tohoto postupu je známý a není zpochybňováný. V posledních letech, díky rozvoji nových technologií, vstupuje stále více do popředí využití možností virtuální či rozšířené reality jako podpůrné metody rehabilitace. Současně se hledají vysvětlení neurofyziologických dějů, na kterých tyto nové techniky pracují.
Jedním ze základních konceptů, které vysvětlují možné fungování těchto metod, je existence tzv. zrcadlových neuronů, Zrcadlové neurony byly poprvé popsány v opičí ventrální premotorické oblasti F5 (6, 22), později též v lobus parietale inferior (8). Existence zrcadlových neuronů u opic je považována za jeden z nejzásadnějších objevů na poli neurofyziologie za posledních několik desítek let. Od té doby bylo provedeno nespočet neinvazivních studií, zkoumajících podobnou existenci i v lidském mozku (9). Zrcadlové neurony jsou specifické asociační neurony, které jsou aktivní jak během provádění určitého pohybu, tak i během pozorování stejného pohybu. Lze předpokládat, že zrcadlové neurony hrají důležitou roli ve schopnosti imitace i u lidské populace (5, 23).
Celý zrcadlový systém se skládá z několika oblastí. Za stěžejní oblast je považován premotorický kortex, dále pak inferiorní frontální kortex, superiorní parietální lalok a zadní část gyrus temporalis superior (23). S objevem a ozřejměním lokalizace a funkce zrcadlových neuronů se začaly rozvíjet výzkumy, které se zaměřují na roli systému zrcadlových neuronů v patofyziologii některých poruch. V roce 1998 přišel neurovědec Ramachandran (21) s prvním úspěšným výzkumem na léčbu fantomových bolestí u amputovaných končetin jedinců. Díky pozitivním výsledkům zrcadlové terapie u fantomových bolestí se rozšířily výzkumy na další klinické jednotky. Nejvýznamnější jsou studie týkající se cévní mozkové příhody (1, 4, 13, 14, 25).
Existence zrcadlových neuronů může být potvrzena několika různými, především neinvazivními, funkčními zobrazovacími metodami mozku, jako například pozitronovou emisní tomografií (PET), funkční magnetickou rezonancí (fMRI), transkraniální magnetickou stimulací (TMS), magnetoencefalografií (MEG) nebo elektroencefalografií (EEG) (7). Přestože funkční magnetická rezonance představuje z hlediska studia intracebrální aktivity nejlepší metodu, nelze ji použít v řadě případů, které se týkají dynamické pohybové aktivity. V těchto případech je EEG metoda velmi vhodnou možností volby. Nedostatkem této rutinní metody je její neschopnost samostatně lokalizovat intracerebrální zdroje elektrické aktivity. Proto se hledaly způsoby, jak tento problém vyšetřit následným postzpracováním získaného elektroencefalografického záznamu. Jedním z možných řešení je použití Standard Low Resolution Brain Electromagnetic Tomography (sLORETA). Tato metoda nabízí efektivní řešení inverzního problému s minimální prostorovou chybou. sLORETA stanovuje distribuci proudové hustoty v celkem 6239 voxelech s prostorovým rozlišením 5x5x5 mm, které pak může zobrazit v 3D Talairachově atlasu (16, 17, 18, 20).
CÍLE A METODY
Cílem naší práce bylo zjistit, zda a jak se liší elektrická mozková aktivita hodnocená pomocí sLORETA zobrazení mezi vlastním aktivním provedením určitého pohybu a pasivním sledováním stejného pohybu, který provádí před probandem terapeut, nebo sledováním stejného pohybu ve 2D videu sledovaného ve virtuálním prostředí.
Studie se zúčastnilo 12 zdravých, vysokoškolských studentů, 8 žen a 4 muži, jejichž průměrný věk byl 24 let. Probandi byli před provedením experimentu poučeni o průběhu měření a podepsali informovaný souhlas. Studie proběhla se souhlasem etické komise UK FTVS pod jednacím číslem 229/2016. Experiment byl proveden v Kineziologické laboratoři UK FTVS.
Elektroencefalografická aktivita byla registrována pomocí telemetrického 32kanálového EEG přístroje Nicolet EEG wireless 32/64 Amplifier firmy Natus Neurology, Inc. (USA). EEG aktivita byla registrována EEG čepicí (Electro-Cap, Inc.) s rozložením elektrod v Mezinárodním systému 10-20, mozková aktivita byla snímána z 19 elektrod: Fp1, Fp2, F7, F3, Fz, F4, F8, T5, T3, C3, Cz, C4, T4, T6, P3, Pz, P4, O1, O2. Vzorkovací frekvence byla 512Hz, pásmová propust 0,5-70Hz, impedanční odpor pod 10kΩ.
Pro sledování pohybu ve virtuálním prostředí bylo vytvořeno 2D video, kde byl zaznamenán pohyb pravou horní končetinou 1. diagonály (flekční i extenční vzorec) dle metody PNF. Tento pohyb prováděl muž, který stal proti bílé zdi, doba trvání byla 2 minuty, pohyb nebyl provázen žádným zvukem Video bylo následně spouštěno prostřednictvím mobilního telefonu Samsung S7 a brýlí pro virtuální realitu Samsung Gear VR.
Měření proběhlo ve dvou dnech, průměrná doba měření jednoho probanda činila 60 minut. Měření probíhalo za standardních podmínek v dopoledních a odpoledních hodinách. Vlastnímu experimentu předcházela 5minutová registrace klidového EEG záznamu probanda vleže na zádech se zavřenýma očima, následně 2 minuty EEG s otevřenýma očima.
Samotný experiment měl 3 části:
- A) aktivní provedení pohybu 1. diagonály z konceptu PNF pravou horní končetinou (ACTIVE),
- B) proband pozoroval terapeuta, který prováděl sám pohyb 1. diagonály dle konceptu PNF pravou horní končetinou (PASSIVE),
- C) proband sledoval ve virtuálním prostředí 2D video s terapeutem, který prováděl identický pohyb 1.diagonály z konceptu PNF pravou horní končetinou (VIDEO).
Každá tato část trvala dvě minuty a probíhala ve stoje. Mezi jednotlivými částmi byla vždy dvouminutová pauza. Během pauzy se proband posadil na lehátko a relaxoval. Po celou dobu provádění všech tří částí byla probandovi snímána elektrická aktivita mozku pomocí EEG přístroje. Pořadí jednotlivých fází bylo randomizováno.
Získaná data byla nejprve zpracována v programu NeuroGuide (Applied Neuroscience, Inc.), kde z každé fáze experimentu byl vybrán 30s bezartefaktový interval. Tento úsek signálu byl vyexportován do sLORETA programu. Následně za použití transformační matrix, zkříženého spektra pro každý subjekt a vybranou frekvenci byly transformovány do sLORETA souborů (16). Výsledné hodnoty představovaly zprůměrněnou zdrojovou intracerebrální aktivitu pro pásmo alfa-1, alfa-2, beta-1 a beta-2 pro všechny probandy. Ke statistickému vyhodnocení byl použit statistický modul, který je součástí programu sLORETA. Pro statistické vyhodnocení byly vybrány sLORETA soubory bez normalizace, s porovnáváním párové skupiny (test A=B), s variantou t-testu s logaritmickou transformací dat s parametrem vyhlazení 0,5 a randomizací dat na hodnotě 5000. Za statisticky významnou hodnotu jsme považovali p≤0,05. Statisticky významnou diferenci zdrojové aktivity pro jednotlivá pásma jsme zobrazili v Talairachově 3D atlasu a výsledky interpretovali pomocí Brodmannových zón.
Porovnávány byly tři situace:
- Aktivní provedení pohybu pravou horní končetinou (ACTIVE) versus klidová EEG aktivita s otevřenýma očima (PRE OE).
- Pasivní sledování pohybu prováděného terapeutem (PASSIVE) versus klidová EEG aktivita s otevřenýma očima (PRE OE).
- Pasivní sledování 2D videa s prováděním pohybu terapeutem (VIDEO) versus klidová EEG aktivita s otevřenýma očima (PRE OE).
VÝSLEDKY
U všech probandů se vyskytovala v průběhu úvodního elektroencefalografického vyšetření alfa aktivita v klidu a při zavřených očích. Tento nález svědčí o schopnosti všech probandů generovat alfa aktivitu i při pohybu a otevřených očích.
ACTIVE versus PRE OE
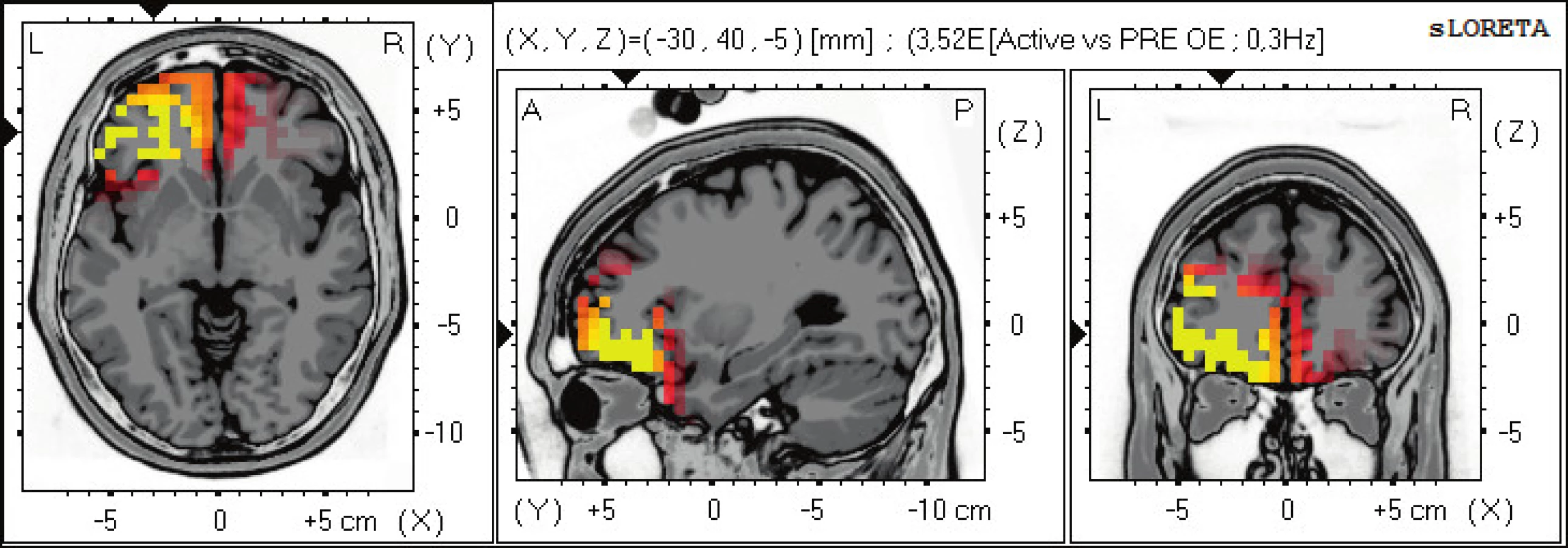
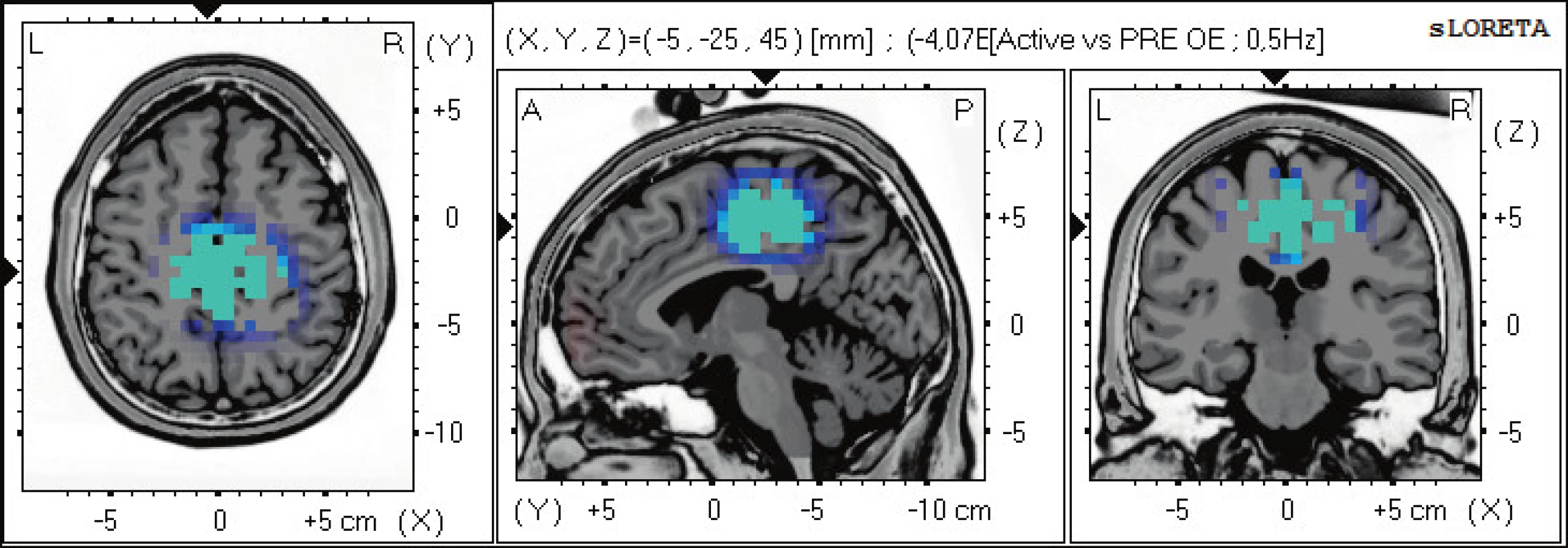
Statisticky signifikantní diference na hladině významnosti p≤0,05 byly zjištěny v pásmu alfa-1, 2 a beta-2. Zvýšená alfa-1 aktivita se vyskytuje ve frontálních oblastech oboustranně v Brodmannových zónách (BAs) 47, 10, 11. Zvýšená alfa-2 aktivita se vyskytuje dominantně nad levou hemisférou v BAs 11, 47, 10 (obr. 1). Snížená aktivita beta-2 se objevuje v BAs limbického systému - gyrus cinguli BAs 31,24, precentrálním gyru BA 6 a precuneu BAs 7,5 (obr. 2).
PASSIVE versus PRE OE
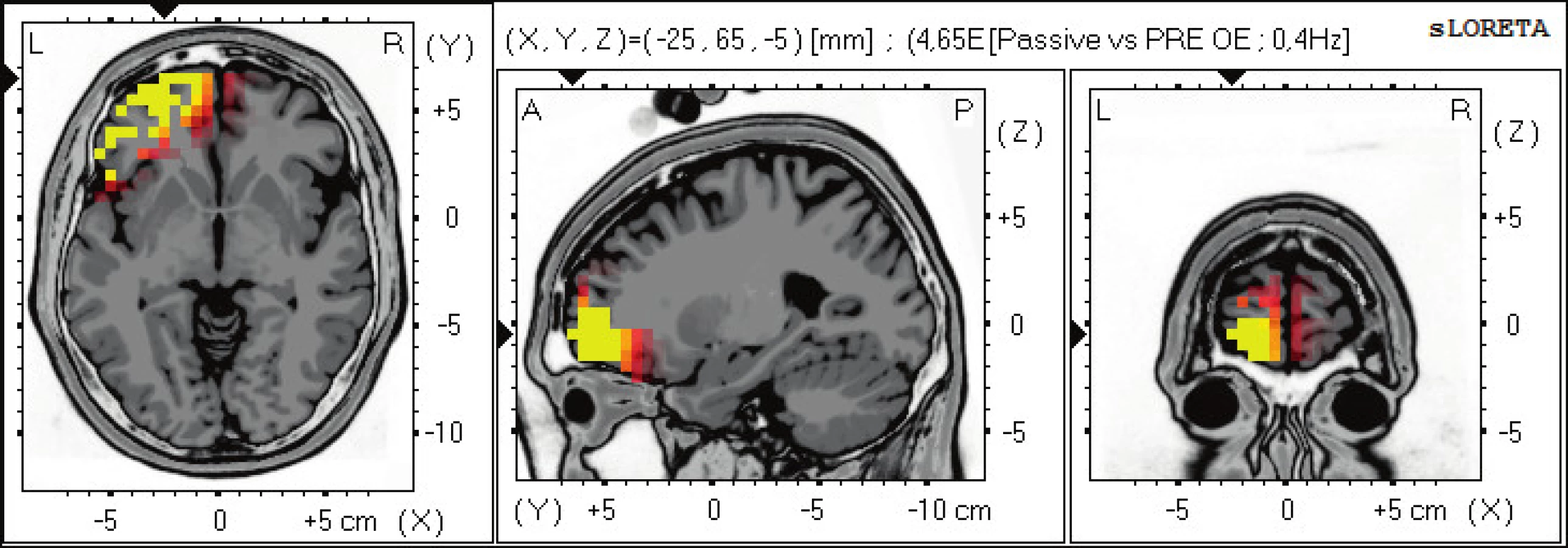
Statisticky signifikantní diference na hladině významnosti p≤0,05 byly zjištěny v pásmu alfa-1, 2 a beta-1, 2. Zvýšená aktivita v alfa-1, 2 se vyskytuje oboustranně ve frontálních oblastech BAs 11, 47, 10 a v gyrus cinguli BA 32. Zvýšená aktivita v beta-1, 2 se objevuje pouze nad levou hemisférou frontálně v BAs 47, 10, 11 (obr. 3).
VIDEO versus PRE OE
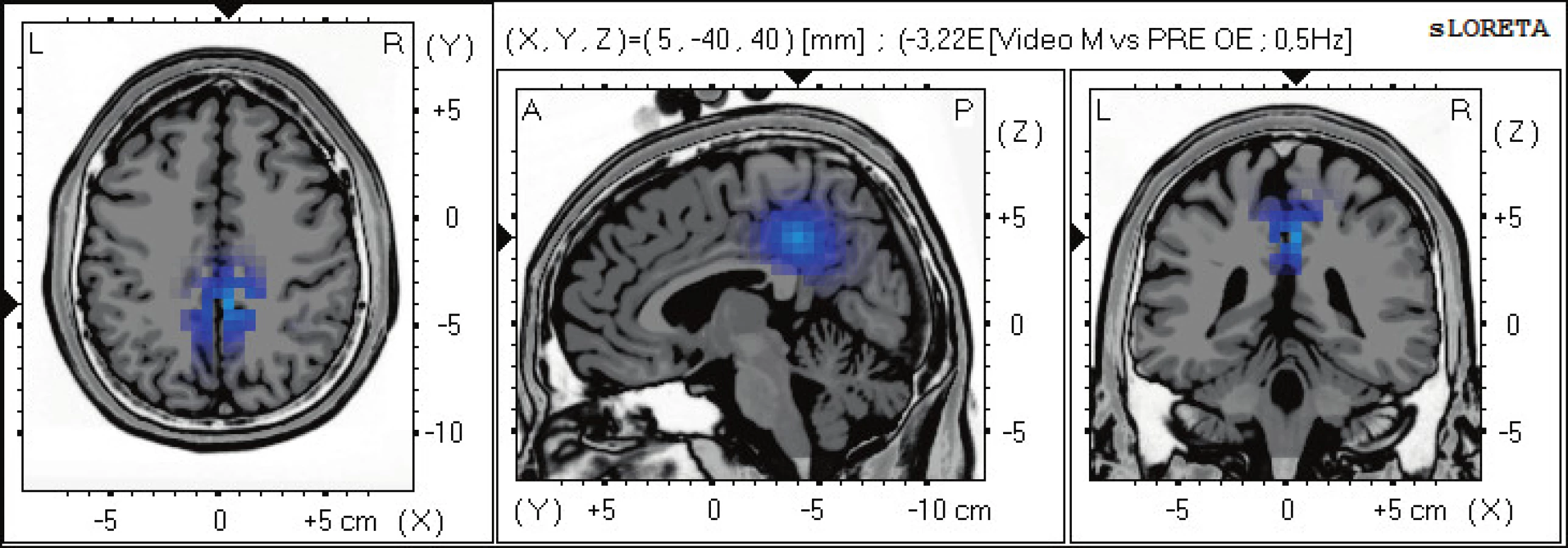
Statisticky signifikantní diference na hladině významnosti p≤0,05 byly pouze v pásmu beta-2, kde se objevuje snížení aktivity v oblasti gyrus cinguli limbického systému BA 31 a precuneu BAs 7,5 (obr. 4)
DISKUSE
Oblasti frontální laloku BAs 10, 11, 46 a 47 jsou součástí dorzolaterálního premotorického laloku, který je spoluzodpovědný za motorické plánování, učení a exekuční funkce, které patří do vyšších kortikálních funkcí. Aktivita v těchto oblasti při provádění pohybu zcela koreluje s jeho probíhající přípravou, kontrolou a eventuálním učením nových pohybových programů (11).
Aktivita v oblasti somatosenzorického asociačního kortexu v parietálním laloku BAs 5,7 je spojena s motorickým učením, prostorovým vnímáním a jeho představivostí a peripersonálním polem. Jeho porucha sje spojena s ideomotorickou apraxií a asterognosí. Aktivita ve frontální premotorické oblast BA 6 je spojena s tvorbou motorických programů a iniciací pohybů. BAs 5,7 a 6 patří do systému zrcadlových neuronů (15). Gyrus cinguli má reciproční spojení s limbickým systémem, ale také s motorickou a premotorickou kůrou. Obě části, přední a zadní cingulární kortex (ACC a PCC), mají silné reciproční spojení s laterálním prefrontálním kortexem (LPFC). Tímto způsobem se spoluúčastní na exekučních funkcích, pracovní paměti, učení a vnějším osobnostním projevu a motorickém plánování (10).
Dorzální cingulární kortex BAs 31, 24 se účastní prostorové navigace a zpracování okolních scén. Současně se spolupodílí na epizodické paměti zahrnující autobiografickou paměť a představu budoucnosti (2, 12).
Při aktivním pohybu nacházíme signifikantní změnu aktivity v oblastech dorzolaterálního prefrontálního kortextu vlevo, somatosenzorického kortexu, premotorické zóny a zadního cingulárního kortexu. Oblast cingulárního kortexu integruje informace z aktuálního postavení těla a okolí, ve kterém se jedinec nachází, tvoří silné spoje mezi asociačními korovými oblasti a prefrontálními oblastmi. Somatosenzorický kortex se účastní ideomotorické činnosti po zpracování vizuoprostorové a proprireceptivní informace a společně s prefrontálními oblastmi provádí další plánování pohybu a přidává osobnostní složku.
Při pasivním pozorování pohybu prováděného terapeutem zůstávají signifikantní změny pouze v oblasti prefrontálního laloku (DLPFC) a cingulárního kortexu. Chybí aktivita v somatosenzorickém a premotorickém kortexu, která může znamenat snížení tvorby vlastní idei pohybu při zachování možnosti pozorovaný program zafixovat v paměti. Objevují se zde oblasti odpovídající okruhu zrcadlových neuronů.
Pozorování 2D videa ve virtuálním prostředí vedlo k vymizení statisticky signifikační aktivity v alfa pásmu, zůstává pouze aktivita v beta-2 pásmu v oblasti zadního cingulárního gyru, který integruje veškeré vjemy z těla a okolí a má spojení s ostatními oblastmi, které se zabývají přípravou a doladěním volního pohybu. Neobjevuje se zde aktivita v prefrontálních oblatech, která by svědčila pro možnosti motorického učení a plánování pohybu. V tomto případě přítomnost aktivity pouze v oblasti zadního cigulárního gyru nejspíše znamená, že při pozorování pohybu dochází k integraci přicházejících vjemů, ale tyto vjemy nemají přímý vliv na motorické učení a plánování. Tedy námi používané video nevyvolalo dostatečnou odezvu v okruhu zrcadlových neuronů, tak jak jsme to viděli při pasivním pozorování pohybu prováděného terapeutem.
ZÁVĚR
Výsledky naší práce poukazují na dvě skutečnosti. První je, že existuje velká podobnost mozkové aktivity při sledování pohybu terapeuta a při vlastním provádění stejného pohybu pacientem. Tento nález zcela koresponduje s existencí zrcadlových neuronů a s výsledky publikovaných studií. Druhým zjištěním je nález diference v mozkové aktivitě mezi sledováním pohybu u reálného terapeuta a sledováním stejného pohybu v 2D videu v brýlích pro virtuální realitu. Ve druhém případě nedochází k aktivitě v prefrontálních oblastech a parietálních oblastech somatosenzorického kortexu. Tento nález nejspíše svědčí o tom, že video nevyvolalo dostatečně silnou odezvu u probandů a nevedlo k aktivaci zrcadlových neuronů, tak jak jsme původně předpokládali. Důvodem nejspíše bude nevhodně zvolený typ videa, včetně použitého prostředí, který nevyvolal dostatečně silnou emoční odezvu, která by mohla vést k odezvě v oblastech motorického řízení. Přesto se domníváme, že současné využití virtuální reality, jako podpůrné rehabilitační metody, může být v budoucnosti běžně používanou metodou. Otázkou dalších výzkumů zůstává, jak má být 2D či 3D video strukturováno, jaké okolní prostředí je potřeba v nahrávaném videu zvolit a zda a jak používat zvukový doprovod.
Tato studie vznikla v rámci Programu PROGRESS na Univerzitě Karlově Q41.
Adresa ke korespondenci:
MUDr. David Pánek, Ph.D.
Katedra fyzioterapie FTVS UK
J. Martího 31
162 52 Praha 6
e-mail: panek@ftvs.cuni.cz
Sources
1. ALTSCHULER, E. L. et al.: Rehabilitation of hemiparesis after stroke with a mirror. The Lancet, 1999, 353.9169, s. 2035-2036.
2. AUGER, S. D., MAGUIRE, E. A.: Assessing the mechanism of response in the retrosplenial cortex of good and poor navigators. Cortex, 49, 2013, 10, s. 2904-2913.
3. BURHANS, L., TALK, A., GABRIEL, M.: Cingulate cortex, editor(s): Neil J. Smelser, Paul B. Baltes, International encyclopedia of the social & Behavioral Sciences, Pergamon, 2001, s. 1799-1805, ISBN 9780080430768.
4. CARVALHO, D. et al.: The mirror neuron system in post-stroke rehabilitation. International Archives of Medicine, 6, 2013, 1, s. 41.
5. CATMUR, C.: Sensorimotor learning and the ontogeny of the mirror neuron system. Neuroscience Letters, 2013, 540, s. 21-27.
6. DI PELLEGRINO, G. et al.: Understanding motor events: a neurophysiological study. Experimental Brain Research, 91, 1992, 1, s. 176-180.
7. FERRARI, P. F., RIZZOLATTI. G.: Mirror neuron research: the past and the future. 2014.
8. FOGASSI, L. et al.: Parietal lobe: from action organization to intention understanding. Science, 2005, 308.5722, s. 662-667.
9. GRAFTON, S. T. et al.: Localization of grasp representations in humans by positron emission tomography. Experimental Brain Research, 112, 1996, 1, s. 103-111.
10. HAYDEN, B. Y., PLATT, M. L.: Cingulate cortex, editor(s): Larry R. Squire, Encyclopedia of Neuroscience, Academic Press, 2009, pp. 887-892, ISBN 9780080450469
11. KOCH, G. et al.: Time course of functional connectivity between dorsal premotor and contralateral motor cortex during movement selection. Journal of Neuroscience, 26, 2006, 28, s. 7452-7459.
12. LEECH, R., SHARP, D. J.: The role of the posterior cingulate cortex in cognition and disease. Brain, 137, 2013, 1, s. 12-32.
13. MICHIELSEN, M. E. et al.: (a) Motor recovery and cortical reorganization after mirror therapy in chronic stroke patients: a phase II randomized controlled trial. Neurorehabilitation and Neural Repair, 25, 2011, .3, s. 223-233.
14. MICHIELSEN, M. E. et al.: (b) The neuronal correlates of mirror therapy: an fMRI study on mirror induced visual illusions in patients with stroke. Journal of Neurology, Neurosurgery & Psychiatry, 2010, jnnp. 2009, s. 194134.
15. OMRANI, M. et al.: Perspectives on classical controversies about the motor cortex. Journal of Neurophysiology, 118, 2017, 3, s. 1828-1848.
16. PÁNEK, D.: Elektroencefalografické koreláty pohybového chování a výkonnostní zátěže. Electroencephalographic correlates of physical movement and behavior and endurance performance. Charles University in Prague, Karolinum Press, 2017.
17. PASCUAL-MARQUI, R. D. et al.: Standardized low-resolution brain electromagnetic tomography (sLORETA): technical details. Methods Find. Exp. Clin. Pharmacol., 2002, 24, Suppl. D, s. 5-12.
18. PASCUAL-MARQUI, R. D. et al.: Low resolution brain electromagnetic tomography (LORETA) functional imaging in acute, neuroleptic-naive, first-episode, productive schizophrenia. Psychiatry Research: Neuroimaging, 90, 1999, 3, s. 169-179.
19. PASCUAL-MARQUI, R. D.: Review of methods for solving the EEG inverse problem. International Journal of Bioelectromagnetism, 1, 1999, 1, s. 75-86.
20. PASCUAL-MARQUI, R. D., MICHEL, C. M., LEHMANN, D.: Low resolution electromagnetic tomography: a new method for localizing electrical activity in the brain. International Journal of Psychophysiology, 18, 1994, 1, s. 49-65.
21. RAMACHANDRAN, V. S.: Encyclopedia of human behavior. Academic Press, 2012.
22. RIZZOLATTI, G. et al.: Premotor cortex and the recognition of motor actions. Cognitive Brain Research, 3, 1996, 3, s. 131-141.
23. RIZZOLATTI, G., CRAIGHERO, L.: The mirror-neuron system. Annu. Rev. Neurosci., 2004, 27, s. 169-192.
24. TALAIRACH, J., TOURNOUX, P.: Co-planar stereotaxic atlas of the human brain: 3-dimensional proportional system: an approach to cerebral imaging. Thieme Medical Publishers, 1988, 122 s. ISBN 3137117011.
25. VAN WIJCK, F. et al.: Making music after stroke: using musical activities to enhance arm function. Annals of the New York Academy of Sciences, 1252, 2012, s. 305-311
Labels
Physiotherapist, university degree Rehabilitation Sports medicineArticle was published in
Rehabilitation & Physical Medicine

2018 Issue 4
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Deep stimulation of the globus pallidus improved clinical symptoms in a patient with refractory parkinsonism and genetic mutation
Most read in this issue
- Zrcadlová terapie a její využití v neurorehabilitaci
- Hodnocení senzorických poruch u dětí
- Vliv dechu na činnost svalů pánevního dna v závislosti na poloze těla
- Fyzioterapia – moderný a dynamický odbor