
Homologs of Retinoblastoma-Associated Protein 46/48 Associate with a Histone Deacetylase to Act Redundantly in Chromatin Silencing
RNA molecules such as small-interfering RNAs (siRNAs) and antisense RNAs (asRNAs) trigger chromatin silencing of target loci. In the model plant Arabidopsis, RNA–triggered chromatin silencing involves repressive histone modifications such as histone deacetylation, histone H3 lysine-9 methylation, and H3 lysine-27 monomethylation. Here, we report that two Arabidopsis homologs of the human histone-binding proteins Retinoblastoma-Associated Protein 46/48 (RbAp46/48), known as MSI4 (or FVE) and MSI5, function in partial redundancy in chromatin silencing of various loci targeted by siRNAs or asRNAs. We show that MSI5 acts in partial redundancy with FVE to silence FLOWERING LOCUS C (FLC), which is a crucial floral repressor subject to asRNA–mediated silencing, FLC homologs, and other loci including transposable and repetitive elements which are targets of siRNA–directed DNA Methylation (RdDM). Both FVE and MSI5 associate with HISTONE DEACETYLASE 6 (HDA6) to form complexes and directly interact with the target loci, leading to histone deacetylation and transcriptional silencing. In addition, these two genes function in de novo CHH (H = A, T, or C) methylation and maintenance of symmetric cytosine methylation (mainly CHG methylation) at endogenous RdDM target loci, and they are also required for establishment of cytosine methylation in the previously unmethylated sequences directed by the RdDM pathway. This reveals an important functional divergence of the plant RbAp46/48 relatives from animal counterparts.
Published in the journal:
Homologs of Retinoblastoma-Associated Protein 46/48 Associate with a Histone Deacetylase to Act Redundantly in Chromatin Silencing. PLoS Genet 7(11): e32767. doi:10.1371/journal.pgen.1002366
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002366
Summary
RNA molecules such as small-interfering RNAs (siRNAs) and antisense RNAs (asRNAs) trigger chromatin silencing of target loci. In the model plant Arabidopsis, RNA–triggered chromatin silencing involves repressive histone modifications such as histone deacetylation, histone H3 lysine-9 methylation, and H3 lysine-27 monomethylation. Here, we report that two Arabidopsis homologs of the human histone-binding proteins Retinoblastoma-Associated Protein 46/48 (RbAp46/48), known as MSI4 (or FVE) and MSI5, function in partial redundancy in chromatin silencing of various loci targeted by siRNAs or asRNAs. We show that MSI5 acts in partial redundancy with FVE to silence FLOWERING LOCUS C (FLC), which is a crucial floral repressor subject to asRNA–mediated silencing, FLC homologs, and other loci including transposable and repetitive elements which are targets of siRNA–directed DNA Methylation (RdDM). Both FVE and MSI5 associate with HISTONE DEACETYLASE 6 (HDA6) to form complexes and directly interact with the target loci, leading to histone deacetylation and transcriptional silencing. In addition, these two genes function in de novo CHH (H = A, T, or C) methylation and maintenance of symmetric cytosine methylation (mainly CHG methylation) at endogenous RdDM target loci, and they are also required for establishment of cytosine methylation in the previously unmethylated sequences directed by the RdDM pathway. This reveals an important functional divergence of the plant RbAp46/48 relatives from animal counterparts.
Introduction
Cytosine DNA methylation is critical for stable silencing of transposable elements (TE) and repetitive sequences and for epigenetic regulation of endogenous gene expression in eukaryotes [1]–[3]. DNA methylation is thought to play an ancestral role in the defense against invasive DNA elements to maintain genome stability and integrity [1]–[3]. In the model plant Arabidopsis, cytosine methylation occurs in three different sequence contexts: CG, CHG and CHH. CG and CHG methylation are heritably maintained respectively by DNA METHYLTRANSFERASE 1 (MET1) and the plant-specific CHROMOMETHYLASE 3 (CMT3). CHH methylation is dynamically maintained through de novo methylation by the DOMAINS-REARRANGED METHYLTRANSFERASE 2 (DRM2) and the RdDM pathway [1].
RdDM is a mechanism by which siRNAs direct de novo cytosine methylation in all sequence contexts of target DNA sequences (complementary to the siRNAs). In Arabidopsis, the plant-specific RNA polymerase Pol IV is thought to initiate silencing by generating single-stranded RNA transcripts that are subsequently converted to double-stranded RNAs (dsRNAs) by RNA-DEPENDENT RNA POLYMERASE 2 (RDR2). dsRNAs are processed by DICER 3 (DCL3) to produce 24-nt siRNAs, which are subsequently loaded to an ARGONAUTE 4 (AGO4)-containing effector complex known as RISC (for RNA-Induced Silencing Complex). Through their interaction with long non-coding RNA transcripts from target loci, generated by the RNA polymerase Pol V, the loaded RISC complexes in association with DRM2 are targeted to RdDM target loci to establish cytosine methylation in CG, CHG and CHH contexts, leading to heterochromatin formation and transcriptional silencing [for reviews, see [2], [4].
siRNAs not only direct DNA methylation, but also trigger repressive histone modifications at RdDM target loci, including histone deacetylation, H3K9 dimethylation (H3K9me2) and H3K27 monomethylation (H3K27me1). Functional loss of the RISC component AGO4 causes a strong reduction in H3K9me2 at the endogenous RdDM target loci including transposable and repetitive elements [5], [6]. Furthermore, it has been shown that at RdDM target loci H3K27 monomethylation, a hallmark for silenced heterochromatin [7], is reduced upon loss of Pol V or AGO4 activity [5]. Together with DNA methylation, these repressive histone modifications establish a silenced heterochromatin state at RdDM target loci.
Histone modifications are involved in the control of DNA methylation. For instance, the H3K9 methyltransferase KRYPTONITE (KYP)/SUVH4 and SUVH4 homologs including SUVH2, SUVH5, SUVH6 and SUVH9, catalyze dimethylation of H3K9, which is recognized and bound by CMT3, leading to the maintenance of CHG methylation [1], [8]. Histone H3 lysine-4 (H3K4) demethylation is also involved in DNA methylation. Recent studies reveal that cytosine methylation is depleted in genomic regions with di- or tri-methylated H3K4 at a genome-wide level [9]; the H3K4 demethylase known as JMJ14/PKDM7B is required for H3K4 demethylation and CHG and CHH methylation at various RdDM target loci [10]. The histone deacetylase HDA6 deacetylates lysines of core histones including H3 and H4, and is required for cytosine methylation in transgenes and silenced rRNA genes [11]–[13]. Multiple genetic screens have revealed that HDA6 is critical for transgene silencing [13], [14]. Loss of HDA6 activity causes a substantial decrease of symmetric cytosine methylation and a moderate reduction in asymmetric CHH methylation in an RdDM-silenced transgene promoter, leading to the transgene reactivation [13]. In addition, disruption of HDA6 function gives rise to histone hyperacetylation and decreased CG and CHG methylation at silenced rRNA gene promoters [11]. HDA6 plays a dual role in silencing of these loci: deacetylating core histones and mediating cytosine methylation [15]. In this way, HDA6 and DNA methylation machinery are thought to work collaboratively to silence target loci.
The histone-binding proteins RbAp46 and RbAp48 are highly homologous WD40-repeat proteins and were first identified in mammalian cells as the tumor-suppressor Rb-binding proteins [16]. Subsequent studies revealed that RbAp46/48 is an integral subunit of multiple chromatin-modifying or -assembly complexes [for a review, see [16]]. RbAp46 forms a complex with the histone acetyltransferase called HAT1 that acetylates H4, whereas RbAp48 is a subunit of the Chromatin Assembly Factor-1 (CAF-1) complex that deposits nucleosomes. Both RbAp46 and RbAp48 are components of several histone deacetylase (HDAC) co-repressor complexes such as the Sin3 complex, which deacetylate core histones to repress target gene expression. In addition, RbAp46/48 is an integral subunit of the evolutionarily conserved Polycomb Repressive Complex 2 (PRC2)-like complexes that catalyze H3K27 trimethylation (H3K27me3), resulting in transcriptional repression. Recent studies have shown that RbAp46/48 functions as a histone (H3–H4 dimer)-binding protein [17]. It is believed that the RbAp46/48-containing complexes interact with histone substrates via either RbAp46 or RbAp48 [17].
RbAp46 and RbAp48 are evolutionarily conserved in animals and plants. There are five homologs in Arabidopsis known as MSI1–MSI5 (MSI for MULTICOPY SUPPRESSOR OF IRA1) [18]. Biological functions of MSI1 and MSI4/FVE have been identified, whereas the functions of MSI2, MSI3 and MSI5 are not known. MSI1 is required for proper vegetative development and plays an essential role in gametophyte and seed development [19], [20]. The MSI1 protein is an integral subunit of the conserved Arabidopsis CAF-1 complex, and has also been found in several PRC2-like complexes [18], [20], [21]. In addition, MSI1 directly interacts with the Arabidopsis Rb homolog (RBR), a key cell-cycle regulator [22], to repress MET1 expression in female gametogenesis, presumably resulting in a reduction in CG methylation [23]. In addition to MSI1, MSI4/FVE also interacts with a plant Rb homolog [24], but the biological implication of this interaction is unclear. FVE has been shown to repress expression of the central floral repressor FLC and several cold-responsive genes in Arabidopsis [24], [25]. FLC inhibits the transition from a vegetative to a reproductive phase (i.e. flowering), and loss of FVE function causes FLC de-repression, resulting in late-flowering [24], [25]. Previous studies reveal that fve mutations cause increased levels of histone acetylation at FLC chromatin [24], [26], indicating that FVE may be involved in deacetylation of FLC chromatin to repress FLC expression. However, recent studies show that loss of FVE function also gives rise to a strong reduction in PRC2-catalyzed H3K27me3, a repressive chromatin mark, in FLC chromatin [27]. Given that the human FVE homologs, RbAp46/48, are subunits of multiple histone-modifying complexes, the mechanisms underlying FVE-mediated transcriptional repression/silencing remain elusive.
FLC plays a crucial role in flowering-time regulation in Arabidopsis and FLC expression is affected by a range of chromatin modifiers (reviewed in refs 28,29). In most rapid-cycling (i.e. early flowering) Arabidopsis ecotypes, FLC expression is repressed or silenced by a group of proteins that mediate or trigger repressive histone modifications at the FLC locus, among which, in addition to FVE, are two conserved RNA 3′end-processing factors called CstF64 and CstF77, RNA-binding proteins known as FCA and FPA, a putative H3K4 demethylase FLOWERING LOCUS D (FLD), and a putative CLF (for CURLY LEAF)-containing PRC2-like complex [for reviews, see [28], [29]]. Furthermore, recent studies show that FLC antisense transcripts trigger FLC silencing [30], [31]. There are two groups of antisense transcripts resulting from alternative polyadenylation. CstF64 and CstF77 function together with FCA and FPA to promote polyadenylation of FLC antisense transcripts at a proximal site, triggering FLC silencing [30], [31]. FLD activity is required for, and acts downstream FCA and FPA in this silencing mechanism [30], [32]. It is believed that the 3′ processing at the proximal polyadenylation site on FLC antisense transcripts leads to co-transcriptional decay of the antisense RNA downstream the proximal site, which may generate aberrant RNAs and trigger repressive histone modifications such as FLD-mediated H3K4 demethylation, and consequent silencing [30].
FLC antisense transcript-triggered silencing is mechanistically different from the siRNA-triggered silencing of RdDM target loci, although both involve RNA molecules. So far, no siRNAs targeting the FLC genomic coding region or 5′ promoter have been detected in Arabidopsis. Consistent with this, knockout of siRNA-silencing pathway components such as Pol IV, Pol V, RDR2 or AGO4 has little effect on FLC silencing [33]. In addition, cytosines in genomic FLC in most Arabidopsis ecotypes are not methylated [34]. Thus, unlike RdDM-mediated silencing, cytosine methylation is not directly involved in FLC silencing. However, both silencing mechanisms require repressive histone modifications such as histone deacetylation, H3K4 demethylation, and/or H3K9 and H3K27 methylation, and involve chromatin silencing.
In this study, we explored the role for FVE and MSI5 in chromatin silencing of various loci targeted by siRNAs or asRNAs. We show that MSI5 acts in partial redundancy with FVE to silence FLC and endogenous RdDM target loci including FWA (containing two tandem repeats), AtMu1 (DNA transposon), AtSN1 (retrotransposon) and IG/LINE (intergenic transcripts). FVE and MSI5 associate with the histone deacetylase HDA6 to form HDAC complexes, and directly interact with the target loci, leading to histone deacetylation and transcriptional silencing. Together, these results show that FVE and MSI5 play an important role in the chromatin silencing of various loci targeted by siRNAs or asRNAs in plants.
Results
MSI5 Functions Redundantly with FVE to Promote the Floral Transition
FVE and MSI5 are Arabidopsis homologs of the human histone-binding RbAp46/48 [18], [24]. The amino acid sequence similarity between FVE and RbAp48 over the entire RbAp48 is 45%, and the similarity between MSI5 and RbAp48 over the entire RbAp48 is also 45%, whereas the identity between FVE and MSI5 over the entire MSI5 is 77% (Figure S1). The high degree of sequence conservation between MSI5 and FVE suggests that these two proteins may have a similar biochemical function.
Previous studies have shown that FVE represses the floral transition in Arabidopsis [24], [25]. We sought to address the biological functions of MSI5. Two loss-of-function mutants of MSI5 carrying insertional T-DNAs were identified, in which the full-length transcription of MSI5 was severely disrupted (Figure 1A and 1B). Grown in long days (LD; 16-hr light/8-hr dark), msi5-1 did not exhibit any visible phenotypes, whereas msi5-2 flowered slightly later than wild-type Col (Figure 1C and 1D), as measured by the developmental criterion of the number of leaves formed prior to flowering, from the primary apical meristem. In short days (8-hr light/16-hr dark), both mutants flowered moderately later than Col (Figure 1E). In both long and short days, msi5-2 flowered later than msi5-1, indicating that msi5-2 is a strong allele. We further confirmed that the moderate late-flowering of msi5-2 was indeed caused by the mutation in a complementation test in which the wild-type copy of MSI5 complemented the msi5-2 mutation (Figure 1F). To examine whether MSI5 acts redundantly with FVE to repress flowering, we introduced both msi5 alleles into fve mutants. In LDs, both msi5-1;fve and msi5-2;fve flowered later than the late-flowering fve mutants (Figure 1D). Of note, msi5-2;fve flowered with 56 leaves on average which is much later than fve (34 leaves on average) (Figure 1D). Hence, MSI5 functions redundantly with FVE to promote Arabidopsis flowering.

Vernalization (an extended period of cold exposure) promotes Arabidopsis flowering. We examined the effect of cold treatment on the flowering times of msi5-1;fve. The late flowering phenotypes of this double mutant were partially suppressed by 7-day cold treatment, and after 35 days of cold exposure, the mutant flowered similar to Col (Figure 1G). It is well known that vernalization largely represses FLC expression to accelerate flowering in Arabidopsis [28], [29]. These data indicate that the late-flowering phenotypes of msi5;fve is largely dependent on FLC and that the activities of MSI5 and FVE are not required for FLC repression by vernalization.
MSI5 Functions in Partial Redundancy with FVE to Repress the Expression of FLC and FLC Homologs
FVE has been shown to repress FLC expression [24], [25]. To examine whether the late flowering of msi5;fve was caused by FLC de-repression, we created an flc;msi5-2;fve triple mutant. In long days, the triple mutant flowered much earlier than msi5-2;fve, but still moderately later than flc (Figure 2A). Hence, the late-flowering of fve;msi5-2 is partly dependent on FLC. We further examined the flowering times of flc, fve;flc and flc;msi5-2;fve in short days, and found that the triple mutant flowered later than fve;flc and flc (Figure 2B), suggesting that FVE and MSI5 may repress other floral repressors to promote flowering, in addition to FLC.
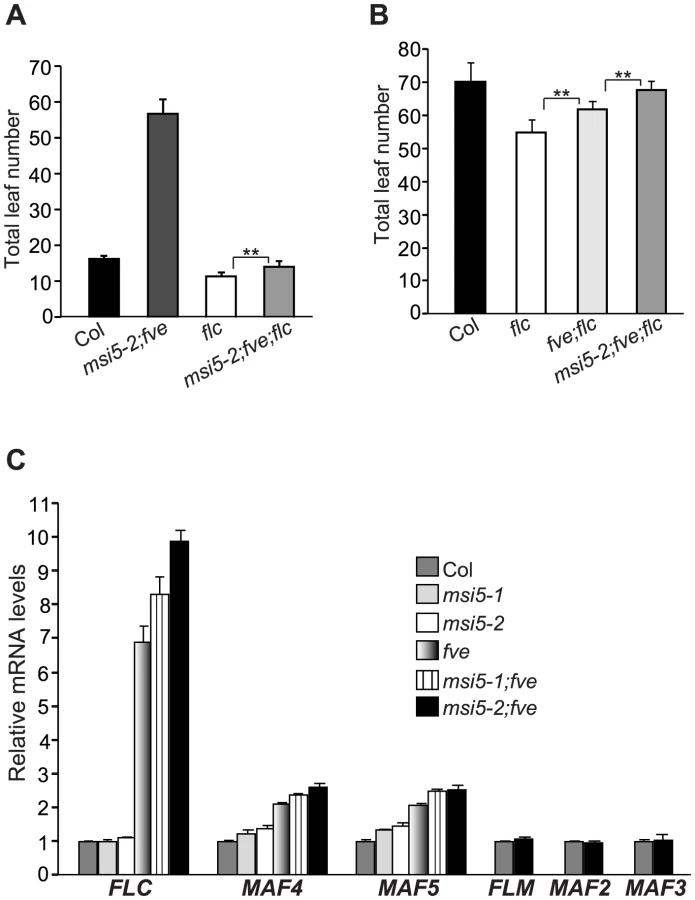
Besides FLC, Arabidopsis has five FLC homologs including FLOWERING LOCUS M (FLM) (also known as MAF1), and MAF2-MAF5 (MAF for MADS BOX AFFECTING FLOWERING); these genes moderately repress flowering [35]–[37]. We quantified transcript levels of FLC and FLC homologs in Col, msi5, fve, and msi5;fve seedlings. FLC expression was slightly increased in msi5-2 compared to Col, whereas it remained unchanged in msi5-1 (Figure 2C). However, both msi5 alleles caused strong increases in FLC transcript levels in the fve background (Figure 2C). Furthermore, we found that in fve mutants both MAF4 and MAF5 were de-repressed, and this de-repression was enhanced upon loss of MSI5 function in the fve background, whereas MAF1, MAF2 and MAF3 expression remained unchanged upon loss of FVE and MSI5 function (Figure 2C). Together, these data show that MSI5 acts redundantly with FVE to repress the expression of MAF4 and MAF5, in addition to FLC, and promote the floral transition.
MSI5 Acts Redundantly with FVE to Silence Endogenous RdDM Target Loci
Recent genetic analyses have revealed that FVE is partly required for proper silencing of the RdDM target loci AtSN1 (retrotransposon) and AtMu1 (DNA transposon), although the underlying mechanism is unknown [38], [39]. This prompted us first to explore whether FVE plays a broad role in silencing of the RdDM target loci including TEs and repetitive elements. We examined the effect of loss of FVE function on the silencing of two other representative RdDM loci, FWA and IG/LINE. FWA, encoding a homeodomain-containing transcription factor that can repress flowering, is sporophytically silenced by cytosine methylation in two sets of tandem repeats containing a sequence related to a SINE (for Short Interspersed Nuclear Element) retroelement located in the 5′ region of FWA [40], [41]. IG/LINE is a spurious intergenic transcript initiated from a flanking solo-LTR (for Long Terminal Repeat) that functions as a promoter [42]. Upon loss of FVE function, FWA and IG/LINE were re-activated in the fve or fve;flc seedlings, respectively (Figure 3A, 3B). We asked whether MSI5 was required for silencing of RdDM target loci. To this end, we first quantified the transcript levels of AtSN1, AtMu1 and IG/LINE in msi5-2;flc and msi5-2;fve;flc seedlings. Both msi5-2 and msi5-2;fve were introduced into the flc background to exclude the possibility that FLC de-repression may affect reactivation of these loci. Loss of MSI5 function alone had little effect on silencing of these three loci; however, upon the combined loss of FVE and MSI5 function, all three loci were strongly re-activated to levels much higher than fve alone (Figure 3B–3D). Next, we measured FWA transcript levels in msi5 and msi5;fve seedlings [in the Col background; note that FLC upregulation does not affect FWA silencing [43]]. FWA is fully silenced in the msi5 seedlings (Figure 3A), but the msi5 mutations greatly enhanced FWA reactivation upon loss of FVE function (Figure 3A), like the situation in the other three loci. Together, these data suggest that MSI5 and FVE may play a broad role in silencing of transposable and repetitive elements in Arabidopsis genome, and that MSI5 functions redundantly with FVE to silence these elements.

These distinct four loci have a common feature, that is, their de novo silencing is established by the siRNA-triggered DNA methylation pathway [40], [42], [44], [45]. To test whether MSI5 and FVE were involved in silencing of TEs other than RdDM target loci, we examined the transcript levels of Ta3 in msi5 and/or fve mutant seedlings (in the flc background), which is a pericentromeric TE that is silenced independently of siRNAs [46]. Loss of MSI5 and/or FVE function did not cause Ta3 reactivation (Figure 3E). These data indicate that MSI5 and FVE may only be required for the silencing of RdDM-targeted TEs and repetitive elements.
MSI5 and FVE Are Required for Both De Novo Asymmetric Cytosine Methylation and Maintenance of Symmetric Methylation at Endogenous RdDM Loci
FWA, AtMu1 and IG/LINE are silenced by cytosine methylation [40]–[42], [44]. We sought to determine whether MSI5 and FVE are required for cytosine methylation in these loci. Using bisulfite sequencing, we examined cytosine methylation at the tandem repeats (TRs), terminal inverted repeats (TIRs) and solo-LTR, respectively, in FWA, AtMu1 and IG/LINE in msi5 and/or fve mutant seedlings (note that these repeats generate siRNAs) (Figure 4A). At the FWA locus, CG methylation was slightly reduced, but a strong reduction in CHG and CHH methylation was observed, in msi5-2;fve compared to wildtype (mCHG: 14% in WT, but 4% in msi5-2;fve; mCHH: 7% in WT, but 2% in msi5-2;fve); neither CHG nor CHH methylation was affected in msi5-2, whereas upon loss of FVE function CHG and CHH methylation was moderately reduced (Figure 4B). At AtMu1, CG methylation was not affected, but CHG methylation was greatly reduced in msi5-2;fve (in the flc background); in addition, CHH methylation was moderately reduced upon loss of FVE and MSI5 function (Figure 4C). At solo-LTR, cytosine methylation in all contexts was reduced upon the combined loss of FVE and MSI5 function (Figure 4D). The reduction of non-CG methylation at the FWA, AtMu1 and solo-LTR loci was further confirmed using the methylation-sensitive restrictive endonucleases Fnu4HI or AluI (Figure S2). Together, these results show that MSI5 and FVE primarily mediate CHH and CHG methylation at RdDM target loci.
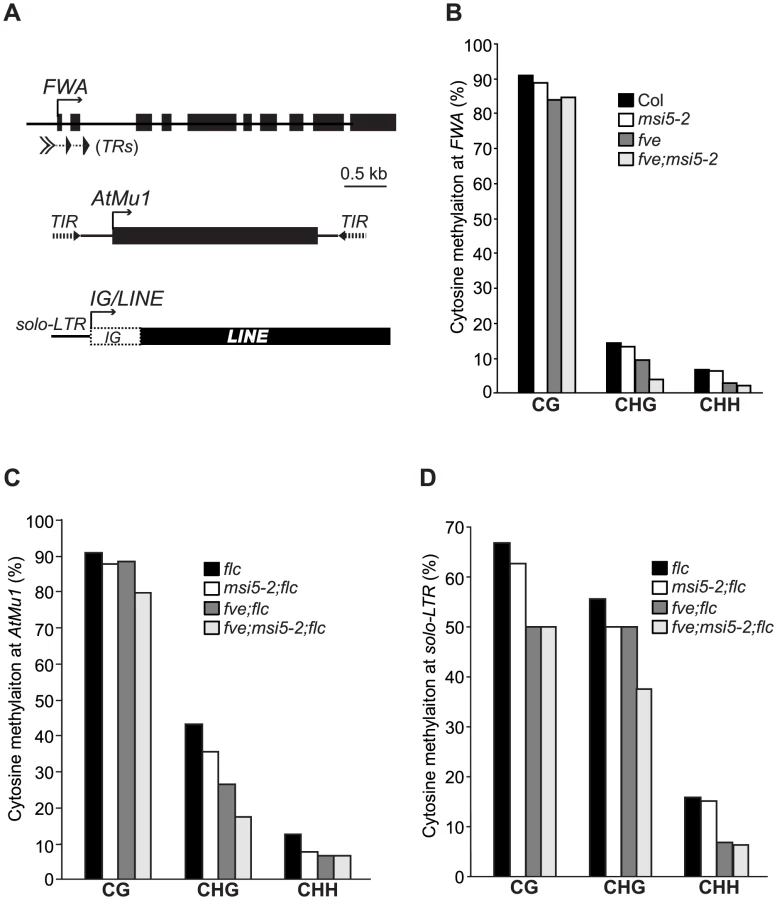
Recent studies show that symmetric CHG methylation is largely maintained by CMT3 in concert with the H3K9 methyltransferase KYP, whereas CHH methylation cannot be maintained, but is de novo methylated by the RdDM pathway [1]. Hence, we conclude that MSI5 and FVE are required for the de novo CHH methylation and maintenance of CHG methylation at the RdDM target loci. Cytosine methylation at both AtMu1 TIRs and solo-LTR causes transcriptional silencing. The reduction in cytosine methylation at the non-transcribed and siRNA-targeted regions (TIRs and solo-LTR) upon loss of FVE and MSI5 function, suggests that these two genes silence RdDM target loci partly by mediating DNA methylation in these loci.
FVE and MSI5 Contribute to the Establishment of DNA Methylation of the FWA Transgene Newly Introduced into Arabidopsis Genome
Both FVE and MSI5 are required for de novo CHH methylation at the endogenous RdDM target loci. We sought to examine whether they could be involved in de novo cytosine methylation in all sequence contexts on previously unmethylated sequences using an FWA transgene assay. When an unmethylated FWA transgene is introduced into Arabidopsis genome, siRNAs from the endogenous FWA are able to target the transgene and through the RdDM pathway direct de novo cytosine methylation in all sequence contexts, leading to its silencing [40], [47]. Otherwise, ectopic FWA expression would give rise to a late-flowering phenotype [47], [48].
We introduced an FWA transgene [47], [48] into flc and flc;msi5-2;fve mutant backgrounds. Consistent with our previous finding that FWA transgene is de novo silenced in the flc background [43], T1 transformants of the flc background flowered only slightly later than flc (Figure 5A). By contrast, T1 transformants of the flc;msi5-2;fve mutant flowered much later than the non-transformed control (Figure 5A). Hence, MSI5 and FVE are required for de-novo silencing of the incoming FWA transgene.
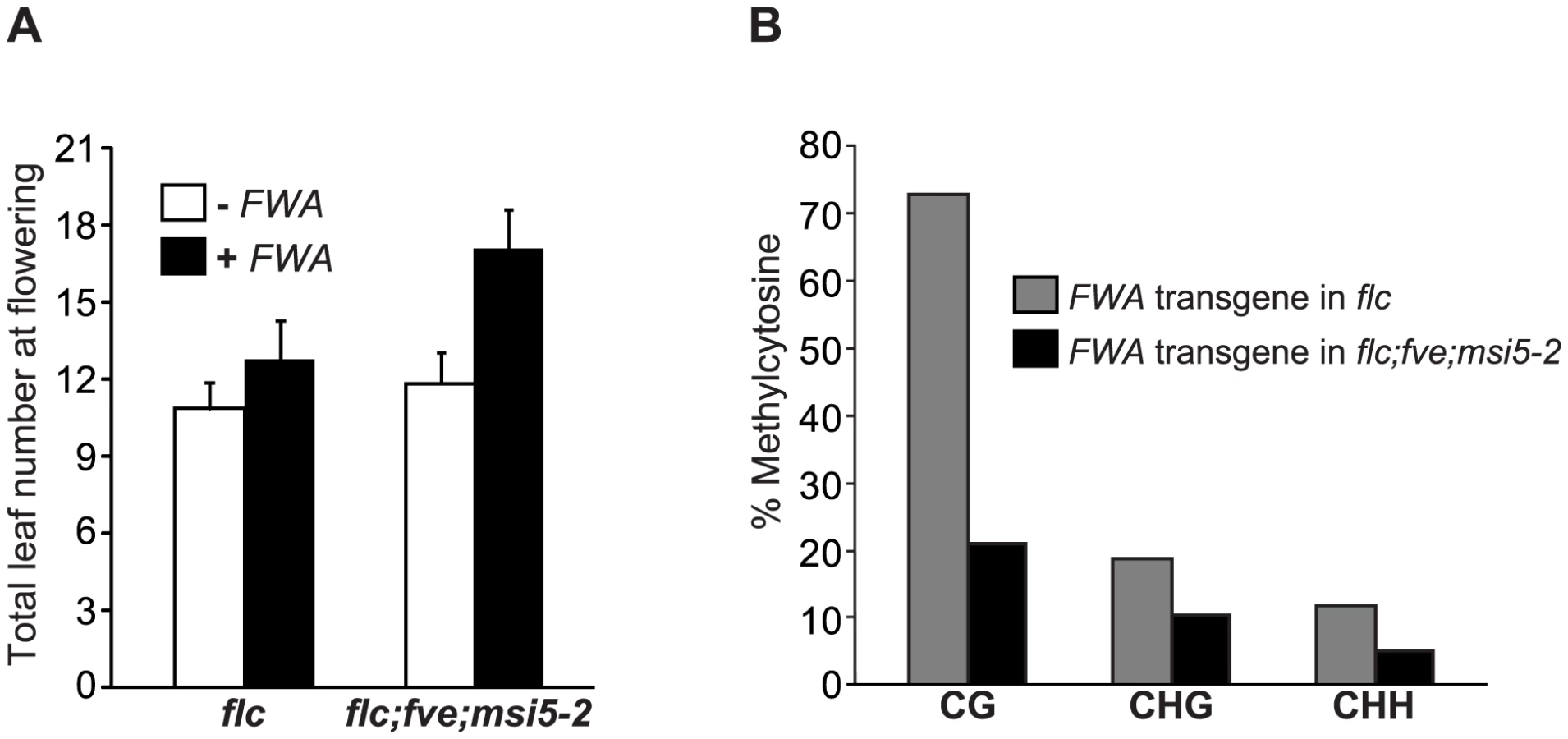
We further examined the methylation state of FWA transgene in the flc and flc;msi5-2;fve backgrounds by bisulphite sequencing, and observed that CG methylation (a primary contributor for FWA silencing), was significantly reduced in the transgene upon the combined loss of FVE and MSI5 function (Figure 5B), in contrast to the slight reduction in CG methylation of the endogenous FWA (Figure 4B). In addition, non-CG methylation of FWA transgene was also reduced in flc;msi5-2;fve compared to the flc background. Together, these results show that FVE and MSI5 are required for the establishment of cytosine methylation in all sequence contexts of the newly introduced FWA transgene and thus de novo FWA silencing.
MSI5 Spatial Expression Pattern Overlaps That of FVE
The functional redundancy of MSI5 with FVE raised the possibility that both genes could be expressed in the same tissues. To test this, we examined the spatial expression patterns of MSI5 and FVE using translational fusions to the reporter gene β-GLUCURONIDASE (GUS); the constructs contained the promoter plus part of the protein-coding region of FVE or MSI5. In seedlings, both MSI5 and FVE were preferentially expressed in shoot apices, root tips and leaf vasculature (Figure 6A–6D). In the reproductive phase, both genes were mainly expressed in styles and the junctions of ovary and receptacle (Figure 6E, 6F). In general, FVE-GUS was expressed at a level higher than that of MSI5-GUS. We confirmed that indeed, FVE transcript levels were much higher than those of MSI5 in both seedlings and floral buds (Figure 6G). This may partly explain why FVE plays a more dominant role in gene silencing than MSI5 does. Given the high protein-sequence homology of MSI5 with FVE, the overlapping expression patterns of these two genes provide an explanation for the functional redundancy of MSI5 with FVE.

Both FVE and MSI5 Associate with HDA6 to Form HDAC Complexes
The mammalian homologs of MSI5 and FVE, RbAp46/48, are subunits of several chromatin-modifying complexes involved in gene silencing such as HDAC co-repressor complexes; RbAp46/48 binds H3–H4 dimers and is thought to recognize and bind histone substrates in these complexes [17]. Using a candidate-gene approach, we explored whether FVE and/or MSI5 could associate with HDA6, an HDAC that has been shown to be involved in FLC repression, DNA methylation maintenance and gene silencing in Arabidopsis [11], [13], [49]. Bimolecular fluorescence complementation (BiFC) [50] was employed to examine whether MSI5 and FVE could associate with HDA6 in plant cells. A non-fluorescent N-terminal EYFP (for Enhanced Yellow Fluorescent Protein) fragment was fused to the full-length FVE and MSI5 individually, whereas a non-fluorescent C-terminal EYFP fragment was fused to the full-length HDA6. nEYFP-FVE and HDA6-cEYFP were simultaneously expressed in onion epidermal cells, and fluorescence was observed in the nuclei, reflecting the physical association of FVE with HDA6 in the nucleus (Figure 7A). Similarly, we also found that MSI5 associated with HDA6 in the nuclei of onion cells (Figure 7B).

Next, we performed protein pull-down assays to confirm the association of HDA6 with FVE and MSI5. Transgenic lines (T3 homozygotes) expressing MSI5-YFP-HA (in msi5-2 background) or FVE-FLAG (in fve background) were created. The MSI5 transgene was fully functional (Figure S3A), whereas the FVE-FLAG was partially functional (Figure S3B. Total proteins were extracted from transgenic seedlings and mixed with the purified GST-HDA6 from E.coli. HDA6 was able to pull down the MSI5 fusion and FVE-FLAG from the protein extracts (Figure 7C–7D). Thus, HDA6 can directly associate with FVE and MSI5 from Arabidopsis seedlings.
We further performed co-immunoprecipitation (co-IP) experiments to determine whether HDA6 is part of a complex with FVE or MSI5 in vivo. First, we created transgenic lines expressing a functional HDA6-FLAG (Figure S3C), and a line expressing a functional HA-FVE (Figure S3D). HDA6-FLAG-expressing plants were crossed to the HA-FVE line or the MSI5-YFP-HA line, and from the resulting F1 seedlings total proteins were extracted for co-IP analysis. Indeed, we found that anti-FLAG (recognizing HDA6-FLAG) immunoprecitated the MSI5 fusion protein and HA-FVE from the seedlings (Figure 7E, 7F). Of note, we detected only a small portion of the HA-FVE protein in the HDA6-FLAG immunoprecipitates from the F1 seedlings (note that no HA-FVE was immunoprecipitated from the seedlings expressing only HA-FVE); this is most likely due to an unstable association of FVE with HDA6. Taken together, these results led us to infer that FVE or MSI5 forms an HDAC complex with HDA6 in Arabidopsis.
HDA6 Is Required for DNA Methylation and Silencing of FVE and MSI5 Target Loci
Consistent with the HDA6 association with FVE and MSI5, recent studies show that HDA6, like FVE and MSI5, represses FLC, MAF4 and MAF5 expression to promote the floral transition [49]. It was of interest therefore to determine whether HDA6 also silences endogenous RdDM target loci. We measured transcript levels of FWA, AtMu1, AtSN1 and IG/LINE in WT (Col) and hda6 seedlings. Indeed, loss of HDA6 activity, like loss of MSI5 and FVE function, caused re-activation of all four loci (Figure 8A–8C). Thus, HDA6, like MSI5 and FVE, is required for silencing of the RdDM target loci.
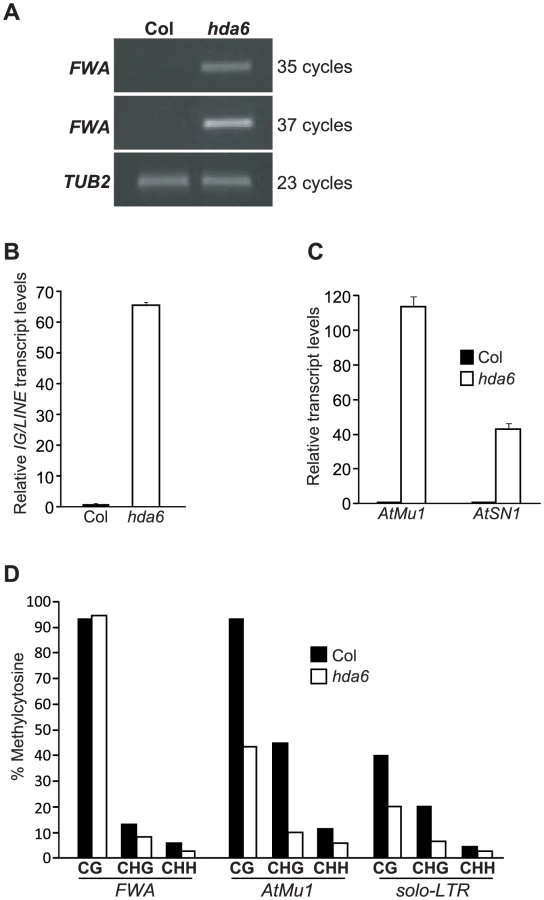
We further examined cytosine methylation state in the FWA, AtMu1 and solo-LTR loci in hda6 seedlings. At FWA, loss of HDA6 function, like of loss of FVE and MSI5 function, caused a reduction in CHG and CHH methylation (Figure 8D). At solo-LTR, cytosine methylation in all sequence contexts was reduced in hda6 compared to wildtype (Figure 8D), similar to the situation in the msi5-2;fve mutant (Figure 4D). In addition, at AtMu1, upon loss of HDA6 function cytosine methylation in all sequence contexts was reduced (Figure 8D). Together, these results show that HDA6, like MSI5 and FVE, is required for cytosine methylation at these RdDM target loci.
FVE Protein Directly Interacts with FLC and RdDM Loci and Is Required for Histone Deacetylation at These Loci
To investigate whether FVE could bind to the chromatin of genes that exhibit altered expression in fve mutants, we performed chromatin immunoprecipitation (ChIP) experiments using the HA-FVE line. Using anti-HA antibodies, we immunoprecipitated DNA fragments from HA-FVE-expressing seedlings (wild-type Col was used as a negative control), and quantified DNA fragments from FLC [the 5′ Intron I region that is essential for FLC silencing; see [26]], AtMu1 (the promoter region immediately downstream the 5′ TIR), solo-LTR and FWA (a region in the silencing tandem repeats) (Figure 9A). Compared with the control, the abundances of HA-FVE protein associated with FLC, solo-LTR and AtMu1 chromatin increased in the HA-FVE line (Figure 9B). In addition, FVE was strongly enriched in FWA (Figure 9B). Of note, the moderate enrichment of HA-FVE at AtMu1 and solo-LTR is likely due to that in the ChIP experiments, anti-HA may not bind effectively to the single HA epitope tag fused to FVE at these TE-containing loci. Taken together, these data suggest that FVE directly interacts with FLC and the three RdDM loci.

FVE or MSI5 forms an HDAC complex with HDA6 and may mediate histone deacetylation for transcriptional silencing. Hence, we examined H3K9 and K14 acetylation state in FVE and MSI5 targets in WT and msi5-2;fve seedlings. H3K9K14 acetylation levels were moderately increased in the AtMu1 promoter region and solo-LTR, and were strongly elevated in the 5′ Intron I of FLC and FWA tandem repeats upon combined loss of FVE and MSI5 function (Figure 9C), consistent with FVE enrichments at these regions. Together, these data suggest that FVE and MSI5 are required for histone deacetylation at FLC and the three RdDM target loci. Given the association of HDA6 with FVE and MSI5, these two proteins may act as part of the HDA6-containing HDAC complexes to mediate deacetylation of target loci and silence their expression.
Discussion
In this study, we show that MSI5 acts redundantly with FVE to silence various loci targeted by siRNAs or asRNAs, including FLC and RdDM target loci. Both FVE and MSI5 form HDAC complexes with HDA6 to mediate histone deacetylation in their target loci. In addition, these two genes function in de novo CHH methylation and maintenance of symmetric cytosine methylation at the endogenous RdDM target loci, and are also required for the establishment of cytosine methylation of the previously unmethylated sequences. Our findings suggest that FVE or MSI5 acts in the context of HDA6-containing co-repressor like complexes to mediate chromatin silencing of developmental genes and RdDM loci of transposable and repetitive elements.
Role for FVE/MSI5-HDA6 in Cytosine Methylation
FVE and MSI5 are required for cytosine methylation at three representative RdDM target loci. These two proteins may mediate cytosine methylation partly via FVE/MSI5-HDA6 complex-catalyzed histone deacetylation. HDA6-catalyzed histone deacetylation is known to be required for cytosine methylation in transgenes and endogenous rRNA genes. It has been shown that silencing of a transgene promoter targeted by RdDM requires HDA6 [13]. The absence of HDA6 activity causes a substantial decrease in symmetric CG and CHG methylation and a moderate decrease in CHH methylation in the promoter region, leading to reactivation of the silenced transgene [13]. Recent studies have shown that HDA6 exhibits a complex interrelationship with cytosine methylation in silenced rRNA genes: loss of HDA6 activity leads to a decrease in symmetric CG and CHG methylation and de-repression of intergenic transcription resulting in overproduction of siRNAs and consequent increase in CHH methylation [11]. Thus, HDA6 is not essentially required for CHH methylation at rRNA genes; however, this does not exclude that HDA6 is still partly involved in this methylation at loci other than rRNAs. A very recent study has revealed that HDA6 is required for both CHH and CHG methylation at a few loci, and partly for CG methylation in some of these loci in Arabidopsis [51].
We have found histone hyperacetylation, loss of cytosine methylation and transcriptional reactivation at the RdDM target loci FWA, AtMu1 and solo-LTR (IG/LINE) upon combined loss of MSI5 and FVE function. This raises the possibility of that the loss of cytosine methylation might result from transcriptional activities at these loci. However, the reasons below argue for that the loss of DNA methylation at least partly causes re-activation of these silent loci. First, both TIRs and solo-LTR are non-transcribed regions. Second, the examined regions with a loss of cytosine methylation including TRs in FWA, TIRs in AtMu1 and solo-LTR in IG/LINE have been shown to generate siRNAs that trigger DNA methylation at these regions leading to transcriptional silencing [40], [42], [44], [45].
In this study, we have revealed that MSI5 and FVE act to silence various loci targeted by siRNAs or asRNAs. This raises the possibility of that these genes could be involved in the production of siRNAs and asRNAs for transcriptional silencing. Recent studies show that loss of FVE function does not affect the production of neither asRNAs from the FLC locus nor the siRNAs from AtMu1, solo-LTR and AtSN1 [30], [38]. We have measured the levels of Pol V-dependent silencing scaffold RNAs from solo-LTR and AtSN1 [5], [45], in mutant seedlings carrying a knockout allele of FVE and/or MSI5, and observed that the loss of FVE and/or MSI5 function does not affect the production of these RNAs (Figure S4). It has also been shown that HDA6 is not involved in siRNA production from the promoter region of a silencing-reporter transgene [13]. Based on these findings, we infer that FVE/MSI5-HDA6 complexes act downstream of or in parallel to siRNA/asRNA production for transcriptional silencing of FLC and RdDM loci. De novo CHH methylation and maintenance of symmetric CHG methylation at the endogenous RdDM loci are catalyzed by the DNA methyltransferases DRM2 and CMT3, respectively, and MSI5/FVE-HDA6 complex-mediated histone deacetylation is expected to facilitate these cytosine methylations. In animals only the CG dinucleotide is methylated [1], hence, the mammalian homologs of FVE and MSI5, RbAp46/48, certainly do not play any roles in CHG and CHH methylation. Our findings on the roles for FVE and MSI5 in cytosine methylation reveal a functional divergence of the plant RbAp46/48 relatives from animal counterparts.
In Arabidopsis, the RdDM pathway controls the establishment of cytosine methylation in the previously unmethylated sequences including CG, CHG and CHH contexts. DRM2 functions as part of the AGO4-containing RdDM effector complex to methylate cytosines in all sequence contexts [52]. Previously, it has been shown that de novo cytosine methylation of the FWA transgene newly introduced into Arabidopsis genome requires the entire RdDM-pathway components including RDR2, DCL3, AGO4, Pol IV and DRM2 [47]. In this study, we have found that FVE and MSI5 are also required for the establishment of cytosine methylation at the incoming FWA. It is likely that the MSI5/FVE-HDA6 complex-mediated histone deacetylation contributes to the establishment of a repressive chromatin environment that promotes DRM2 to catalyze cytosine methylation at the previously unmethylated sequences.
FVE/MSI5-HDA6 Co-Repressor–Like Complexes
The mammalian FVE and MSI5 homologs, RbAp46/48, are subunits of multiple chromatin-modifying complexes such as hHAT1 (involved in gene activation), PRC2 and HDAC co-repressor complexes [16]. So far, FVE and MSI5 have been found to be involved only in transcriptional silencing. Previous studies have shown that loss of FVE function causes reduced H3K27me3 and histone hyperacetylation at the FLC locus, indicating that FVE might act in the context of a PRC2-like complex and/or an HDAC co-repressor-like complex for FLC silencing under normal growth conditions [24], [26], [27]. A very recent study suggests that FVE may be part of a CLF-containing PRC2-like complex to deposit H3K27me3 in FLC and silence its expression [53]; however, the findings described below argue against this notion. Firstly, a gain-of-function clf allele clf-59 [27], suppresses FLC expression in the msi5-2;fve double mutant, resulting in early flowering (Figure S5A); hence, CLF functions independently of FVE and MSI5 to regulate FLC expression. Secondly, to genetically test whether FVE could be part of a CLF-PRC2 complex, we created a clf;fve double mutant in which both clf (clf-29) and fve (fve-4) are null loss-of-function alleles [19], [24], examined FLC de-repression in clf, fve and clf;fve seedlings, and found that CLF and FVE act synergistically to silence FLC expression (Figure S5B), suggesting that FVE may not be part of the CLF complex. Thirdly, in this study we found that FVE and MSI5 silence RdDM target loci, which typically lack of H3K27me3 deposited by PRC2 complexes [54]. Lastly, we carried out co-IP experiments to determine whether FVE and CLF could be in a complex using F1 seedlings expressing a fully-functional GFP-CLF [55] and the HA-FVE fusion, but did not detected an association of CLF with FVE in seedlings (Figure S6). Together, these findings suggest that FVE and MSI5, unlike RbAp46/48, may not act as part of PRC2-like complexes to silence target-locus expression. At the FLC locus, both FVE/MSI5-HDA6 and CLF-PRC2 complexes directly repress its expression [56], and may act in concert to establish a repressive chromatin environment at FLC for its transcriptional silencing (see Figure 10 as described next).

In mammals, RbAp46/48 is an integral subunit of Class I HDAC co-repressor complexes [57], [58], and several of these complexes such as the BRAF-HDAC complex contain the H3K4 demethylase Lysine-Specific Demethylase 1 (LSD1) [59], a mammalian homolog of the Arabidopsis FLD [60]. HDA6, like the Class I HDACs, is an RPD3 (for Reduced Potassium Deficiency 3)-type histone deacetylase [13]. A recent study has revealed that HDA6 forms a complex with FLD to repress FLC expression and promote flowering [61]. In this study, we have found that HDA6 also forms a complex with FVE or MSI5. Together, these findings led us to infer that HDA6 and FLD form an HDAC co-repressor like complex with FVE or MSI5. Consistent with this, in a co-IP experiment using a line expressing a fully functional FLD-myc [62] and the MSI5-YFP-HA fusion, we have confirmed that indeed FLD is in a complex with MSI5 in Arabidopsis seedlings (Figure S7). Furthermore, like MSI5, FVE and HDA6, FLD is also required for the silencing of FLC, FLC homologs and the RdDM target loci including AtMu1, AtSN1 and IG/LINE [38], [61]. Taken together, these Arabidopsis homologs of the mammalian Class I HDAC co-repressor complex components may form HDAC co-repressor like complexes to silence developmental genes and TEs.
The loss of HDA6 function appears to cause a greater reactivation of AtMu1 than that upon the combined loss of FVE and MSI5 function, indicating that HDA6 silences this locus partly independent of MSI5 and FVE. One explanation is that MSI1, MSI2, and/or MSI3 may also participate in HDA6-mediated silencing. HDA6 plays multiple roles in Arabidopsis development. In addition to the acceleration of floral transition, HDA6 is also involved in plant senescence and acts redundantly with its homolog HDA19 to repress embryonic traits in vegetative growth [49], [63], [64], in which FVE/MSI5 appears not to be involved (data not shown). These observations indicate that HDA6 may act partially independent of FVE and MSI5 to silence developmental genes in Arabidopsis.
Models for the Functions of MSI5 and FVE in Chromatin Silencing
MSI5 acts redundantly with FVE to silence the developmental gene FLC and RdDM loci targeted by the silencing triggers asRNAs or siRNAs, respectively. Transcriptional silencing of these loci requires repressive chromatin modifications including histone deacetylation, H3K4 demethylation, H3K9 methylation, and/or H3K27 methylation.
At the FLC locus, the transcriptional silencing is triggered by FLC antisense transcripts and/or aberrant RNA molecules derived from the 3′ processing of asRNAs [30]. In the chromatin silencing at FLC, the targeted 3′ processing of FLC antisense transcripts produces RNA triggers that lead to the recruitment of repressive histone-modification activities on FLC chromatin. As noted above, FLD and HDA6 form a co-repressor like complex with FVE or MSI5. Consistent with this, FLD is required for both H3K4 demethylation and histone deacetylation on FLC chromatin [26], [32], [43]. Moreover, the loss-of-function fld and fve mutations act largely non-additively to delay the floral transition (caused by FLC de-repression) [38]. As illustrated in Figure 10A, it is very likely that the RNA molecules may trigger the recruitment of FLD-FVE/MSI5-HDA6 complexes to FLC chromatin, resulting in repressive histone deacetylation and H3K4 demethylation. In addition, the HDA6 complexes are expected to act collaboratively with the CLF-PRC2 complex that deposits repressive H3K27me3 at FLC. Together, these histone modifiers establish a repressive chromatin environment at the FLC locus leading to heterochromatin-like formation and consequent FLC silencing (Figure 10A).
At the RdDM target loci of transposable and repetitive elements, both cytosine methylation and repressive chromatin modifications such as histone deacetylation, contribute to transcriptional silencing. For instance, FWA silencing is typically caused by symmetric CG methylation [40], [41]. We have found that loss of FVE and MSI5 function leads to histone H3 hyperacetylation at the endogenous FWA and ectopic FWA activation in sporocytes, but only a slight reduction in CG methylation. This suggests that MSI5/FVE-HDA6-mediated histone deacetylation plays a direct role in FWA silencing, in addition to promoting CHG and CHH methylation at this locus. In the chromatin silencing at the RdDM target loci (Figure 10B), MSI5/FVE-containing complexes mediate histone deacetylation and possibly, H3K4 demethylation, on one hand, directly represses target locus expression, and on the other hand, together with H3K9 dimethylation and/or H3K27 monomethylation, establish a repressive chromatin environment that promotes cytosine methylation (mainly CHG and CHH methylation), which may reinforce the repressive histone modifications. Together, these modifications lead to silent heterochromatin formation and consequent transcriptional silencing. FLD, the putative H3K4 demethylase, may act as part of the FVE/MSI5-HDA6 complexes to silence some of the RdDM loci such as AtMu1, IG/LINE and AtSN1 because FLD has been shown to be required for silencing of these loci. Previously we have observed that FLD appears not to be required for FWA cytosine methylation and silencing, but two FLD homologs known as LDL1 and LDL2 mediate FWA silencing [43]. It is likely that HDA6 and FLD may form a co-repressor like complex with FVE or MSI5 to silence certain RdDM target loci, whereas at some other loci, HDA6 and FVE/MSI5 may form a complex with other components for transcriptional silencing.
Materials and Methods
Plant Materials and Growth Conditions
Arabidopsis thaliana fve-4 [24], flc-3 [65], hda6/axe1-5 [49], clf-29 [19] and clf-59 [27] were described previously. The msi5-1 (Salk_004926) and msi5-2 (Salk_116714) alleles were isolated from the SALK collection [66]. Plants were grown under cool white fluorescent lights in long days (16-hr light/8-hr dark) or short days (8-hr light/16-hr dark).
Bimolecular Fluorescence Complementation Assay
The full-length coding sequences for HDA6, FVE and MSI5 were translationally fused with either an N-terminal EYFP fragment in the pSAT1A-nEYFP-N1/pSAT1-nEYFP-C1 vectors and/or a C-terminal EYFP fragment in the pSAT1A-cEYFP-N1/pSAT1-cEYFP-C1-B vectors (www.bio.purdue.edu/people/faculty/gelvin/nsf/index.htm). Using the Helium biolistic gene transformation system (Bio-Rad), onion epidermal cells were transiently co-transformed by appropriate plasmid pairs as indicated in Figure 7. EYFP fluorescence in the onion cells was observed and imaged using a Zeiss LSM 5 EXCITER upright laser scanning confocal microscopy (Zeiss) within 24–48 hrs after bombardment.
Plasmid Constructions
To create HA-FVE fusion, the full-length FVE coding sequence (1.5 kb) was first cloned into the entry vector pENTR4 (Invitrogen), and subsequently, the FVE fragment was inserted downstream of the 35S promoter and the single HA epitope in the pEarlyGate 201 vector [67] via gateway technology (Invitrogen), resulting in the p35S-HA-FVE plasmid. For MSI5-YFP-HA construction, the full-length MSI5 coding sequence (1.5 kb) was inserted downstream of the 35S promoter, but upstream of YFP followed by the single HA epitope in the pEarlyGate 101 vector [67], resulting in the p35S-MSI5-YFP-HA plasmid. To construct GST-HDA6, the full-length FVE coding sequence was cloned into downstream of GST in the protein expression vector pGEX-4T-1. To construct FVE-GUS, a 4,073-bp FVE genomic fragment (from −1,886 to +2,187; A of the start codon as +1) including a 1.9-kb native promoter plus a 2.2-kb genomic coding region was inserted upstream of the GUS reporter gene in the pMDC162 vector [68]; the genomic coding sequence was in frame with GUS. For MSI5-GUS construction, we inserted a 2,145-bp MSI5 genomic fragment (from −438 to +1,707) into upstream of the GUS reporter gene in pMDC162; the genomic coding sequence of MSI5 was in frame with GUS. For the construction of a binary plasmid harboring a wild-type copy of MSI5, a 5.1-kb genomic fragment including the 5′ promoter (1.6 kb), genomic coding sequence (3.2 kb) and 3′ end (0.3 kb), was cloned into pBGW [69]. To clone the gain-of-function clf-59 allele with a single point mutation [27], a 6.5-kb genomic fragment of clf-59 consisting of a 1.3-kb 5′ promoter, 4.5-kb genomic coding sequence and 0.7-kb 3′ end, was amplified from a Ws background and cloned into the binary vector pHGW [69].
Real-Time Quantitative RT-PCR
Total RNAs were extracted from aerial parts of 10-d-old seedlings grown in long days as described previously [43]. The total RNAs were subsequently treated with ‘TURBO DNA-Free’ (Ambion) to remove residual genomic DNA. After reverse transcription, the real-time quantitative PCR was carried out on an ABI Prism 7900HT sequence detection system as previously described [43]. Primers used to amplify the cDNAs of FLC, FLM, MAF2-5, IG/LINE, and TUB2 (At_5g62690) have been previously described [38], [70]. The primer pairs used for MSI5, FVE, FWA, AtMu1, AtSN1 and Ta3 amplification are specified in Table S1. Each sample was quantified in triplicate and normalized to the endogenous control TUB2. Bars indicate standard deviations of triplicate measurements.
Bisulfite Genomic Sequencing
DNA was extracted from 10-d-old seedlings grown in long days, and subsequently, approximately 0.2-µg genomic DNA from each genotype was treated with bisulfite using the EpiTect Bisulfite kit (Qiagen) according to the manufacturer's instruction. The bottom strands of the endogenous FWA (tandem-repeat region), solo-LTR and AtMu1 (the 3′ terminal-inverted-repeat region) were amplified by PCR, and cloned into the T-Easy vector (Promega). The primers used for the endogenous FWA amplification has been described previously [43], and the primer pairs for solo-LTR and AtMu1 amplification are specified in Table S1. Analysis of cytosine methylaion of the FWA transgene in T1 transformants of flc and flc;msi5;fve was performed as described previously [40], [43].
Protein Pull-Down Assay
Total proteins were extracted from 10-d old seedlings. Briefly, 0.5-g seedlings were ground in liquid nitrogen and homogenized in 1.0-ml extraction buffer (50 mM Tris-HCl pH 7.4, 100 mM NaCl, 10% glycerol, 0.1% NP-40, 1.0 mM PMSF) supplemented with 1× Roche protease inhibitor (without EDTA). Subsequently, the GST-HDA6 or GST proteins affinity-purified from the E.coli strain BL21 (DE3) together with the glutathione-linked resins (Sigma) were added into 1.0-ml protein extracts and incubated for 4 hrs at 4°C. The protein pull-downs were analyzed by immunoblotting using anti-HA (Roche, Cat#: 12-013-819-001) or anti-FLAG (Sigma, Cat#: A8592).
Co-Immunoprecipitation
Immunoprecipitation experiments were performed as described previously [71]. Briefly, 0.4-g seedlings were harvested and ground in liquid nitrogen, and subsequently, total proteins were extracted and immunoprecipitated with anti-FLAG M2 affinity gel (Sigma, Cat#: A2220). Proteins in the immunoprecipitates were detected by western blotting with anti-FLAG (Sigma, Cat#: A8592) or anti-HA (Roche, Cat#: 12-013-819-001).
ChIP-Quantitative PCR
ChIP experiments were performed with 10-d-old seedlings largely as previously described [72], [73]. Briefly, nucleus fraction was isolated, and subsequently, immunoprecipitations were carried out using the polyclonal anti-acetylated histone H3 (Lys 9 and Lys 14) (Millipore, Cat#: 06-599B) or anti-HA (Sigma, Cat#: H6908). Quantitative measurements of genomic fragments of FLC, AtMu1, solo-LTR, MAF3 and TUB2 (as the internal normalization control) were performed using SYBR Green PCR master mix (Applied Biosystems). Quantitative measurements of FWA genomic regions and ACTIN2 (At_3g18780; served as the internal normalization control for FWA enrichment) were performed on an ABI Prism 7900HT sequence detection system using TaqMan MGB probes (FAM dye–labeled) as described previously [43]. Each ChIP sample was quantified in triplicate. The primers used to amplify FLC and TUB2 were described previously [43], and the primer pairs for AtMu1, solo-LTR and MAF3 amplification are specified in Table S1. Rationale for calculation of the fold enrichment of HA-FVE in the HA-FVE line over the control line (Col) is as follows: in the HA-FVE line a gene of interest (eg. FLC) was first normalized to TUB2 or ACTIN2, and in the control line the gene of interest was similarly normalized; subsequently, the normalized value from the HA-FVE line was further normalized by the value from the control line to obtain a value of fold enrichment for the gene of interest. A similar rationale was adopted for the calculation of fold enrichment of acetylated H3 in msi5-2;fve over Col.
Supporting Information
Zdroje
1. LawJAJacobsenSE 2010 Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet 11 204 220
2. MatzkeMKannoTDaxingerLHuettelBMatzkeAJ 2009 RNA-mediated chromatin-based silencing in plants. Curr Opin Cell Biol 21 367 376
3. SazeHKakutaniT 2010 Differentiation of epigenetic modifications between transposons and genes. Curr Opin Plant Biol 14 81 87
4. ChinnusamyVZhuJK 2009 RNA-directed DNA methylation and demethylation in plants. Sci China C Life Sci 52 331 343
5. WierzbickiATReamTSHaagJRPikaardCS 2009 RNA polymerase V transcription guides ARGONAUTE4 to chromatin. Nat Genet 41 630 634
6. ZilbermanDCaoXJacobsenSE 2003 ARGONAUTE4 control of locus-specific siRNA accumulation and DNA and histone methylation. Science 299 716 719
7. JacobYStroudHLeblancCFengSZhuoL 2010 Regulation of heterochromatic DNA replication by histone H3 lysine 27 methyltransferases. Nature 466 987 991
8. EbbsMLBenderJ 2006 Locus-specific control of DNA methylation by the Arabidopsis SUVH5 histone methyltransferase. Plant Cell 18 1166 1176
9. ZhangXBernatavichuteYVCokusSPellegriniMJacobsenSE 2009 Genome-wide analysis of mono-, di- and trimethylation of histone H3 lysine 4 in Arabidopsis thaliana. Genome Biol 10 R62
10. DelerisAGreenbergMVAusinILawRWMoissiardG 2010 Involvement of a Jumonji-C domain-containing histone demethylase in DRM2-mediated maintenance of DNA methylation. EMBO Rep 11 950 955
11. EarleyKWPontvianneFWierzbickiATBlevinsTTuckerS 2010 Mechanisms of HDA6-mediated rRNA gene silencing: suppression of intergenic Pol II transcription and differential effects on maintenance versus siRNA-directed cytosine methylation. Genes Dev 24 1119 1132
12. HeXJHsuYFPontesOZhuJLuJ 2009 NRPD4, a protein related to the RPB4 subunit of RNA polymerase II, is a component of RNA polymerases IV and V and is required for RNA-directed DNA methylation. Genes Dev 23 318 330
13. AufsatzWMetteMFvan der WindenJMatzkeMMatzkeAJ 2002 HDA6, a putative histone deacetylase needed to enhance DNA methylation induced by double-stranded RNA. Embo J 21 6832 6841
14. ProbstAVFagardMProuxFMourrainPBoutetS 2004 Arabidopsis HISTONE DEACETYLASE HDA6 is required for maintenance of transcriptional gene silencing and determines nuclear organization of rDNA repeats. Plant Cell 16 1021 1034
15. AufsatzWStoiberTRakicBNaumannK 2007 Arabidopsis HISTONE DEACETYLASE 6: a green link to RNA silencing. Oncogene 26 5477 5488
16. LoyolaAAlmouzniG 2004 Histone chaperones, a supporting role in the limelight. Biochim Biophys Acta 1677 3 11
17. MurzinaNVPeiXYZhangWSparkesMVicente-GarciaJ 2008 Structural basis for the recognition of histone H4 by the histone-chaperone RbAp46. Structure 16 1077 1085
18. HennigLBouveretRGruissemW 2005 MSI1-like proteins: an escort service for chromatin assembly and remodeling complexes. Trends Cell Biol 15 295 302
19. BouveretRSchonrockNGruissemWHennigL 2006 Regulation of flowering time by Arabidopsis MSI1. Development 133 1693 1702
20. KohlerCHennigLBouveretRGheyselinckJGrossniklausU 2003 Arabidopsis MSI1 is a component of the MEA/FIE Polycomb group complex and required for seed development. Embo J 22 4804 4814
21. De LuciaFCrevillenPJonesAMGrebTDeanC 2008 A PHD-Polycomb repressive complex 2 triggers the epigenetic silencing of FLC during vernalization. Proc Natl Acad Sci U S A 105 16831 16836
22. BorghiLGutzatRFuttererJLaizetYHennigL 2010 Arabidopsis RETINOBLASTOMA-RELATED is required for stem cell maintenance, cell differentiation, and lateral organ production. Plant Cell 22 1792 1811
23. JullienPEMosqunaAIngouffMSakataTOhadN 2008 Retinoblastoma and its binding partner MSI1 control imprinting in Arabidopsis. PLoS Biol 6 e194
24. AusinIAlonso-BlancoCJarilloJARuiz-GarciaLMartinez-ZapaterJM 2004 Regulation of flowering time by FVE, a retinoblastoma-associated protein. Nat Genet 36 162 166
25. KimHJHyunYParkJYParkMJParkMK 2004 A genetic link between cold responses and flowering time through FVE in Arabidopsis thaliana. Nat Genet 36 167 171
26. HeYMichaelsSDAmasinoRM 2003 Regulation of flowering time by histone acetylation in Arabidopsis. Science 302 1751 1754
27. DoyleMRAmasinoRM 2009 A single amino acid change in the Enhancer of Zeste ortholog CURLY LEAF results in vernalization-independent, rapid flowering in Arabidopsis. Plant Physiol 151 1688 1697
28. AmasinoR 2010 Seasonal and developmental timing of flowering. Plant J 61 1001 1013
29. CrevillenPDeanC 2011 Regulation of the floral repressor gene FLC: the complexity of transcription in a chromatin context. Curr Opin Plant Biol 14 38 44
30. LiuFMarquardtSListerCSwiezewskiSDeanC 2010 Targeted 3′ processing of antisense transcripts triggers Arabidopsis FLC chromatin silencing. Science 327 94 97
31. HornyikCTerziLCSimpsonGG 2010 The Spen family protein FPA controls alternative cleavage and polyadenylation of RNA. Dev Cell 18 203 213
32. LiuFQuesadaVCrevillenPBaurleISwiezewskiS 2007 The Arabidopsis RNA-binding protein FCA requires a Lysine-Specific Demethylase 1 homolog to downregulate FLC. Mol Cell 28 398 407
33. SwiezewskiSCrevillenPLiuFEckerJRJerzmanowskiA 2007 Small RNA-mediated chromatin silencing directed to the 3′ region of the Arabidopsis gene encoding the developmental regulator, FLC. Proc Natl Acad Sci U S A 104 3633 3638
34. FinneganEJKovacKAJaligotESheldonCCJames PeacockW 2005 The downregulation of FLOWERING LOCUS C (FLC) expression in plants with low levels of DNA methylation and by vernalization occurs by distinct mechanisms. Plant J 44 420 432
35. ScortecciKCMichaelsSDAmasinoRM 2001 Identification of a MADS-box gene, FLOWERING LOCUS M, that represses flowering. Plant J 26 229 236
36. RatcliffeOJKumimotoRWWongBJRiechmannJL 2003 Analysis of the Arabidopsis MADS AFFECTING FLOWERING gene family: MAF2 prevents vernalization by short periods of cold. Plant Cell 15 1159 1169
37. KimDHSungS 2010 The Plant Homeo Domain finger protein, VIN3-LIKE 2, is necessary for photoperiod-mediated epigenetic regulation of the floral repressor, MAF5. Proc Natl Acad Sci U S A 107 17029 17034
38. BaurleIDeanC 2008 Differential interactions of the autonomous pathway RRM proteins and chromatin regulators in the silencing of Arabidopsis targets. PLoS ONE 3 e2733
39. VeleyKMMichaelsSD 2008 Functional redundancy and new roles for genes of the autonomous floral-promotion pathway. Plant Physiol 147 682 695
40. ChanSWZhangXBernatavichuteYVJacobsenSE 2006 Two-step recruitment of RNA-directed DNA methylation to tandem repeats. PLoS Biol 4 e363
41. KinoshitaYSazeHKinoshitaTMiuraASoppeWJ 2007 Control of FWA gene silencing in Arabidopsis thaliana by SINE-related direct repeats. Plant J 49 38 45
42. HuettelBKannoTDaxingerLAufsatzWMatzkeAJ 2006 Endogenous targets of RNA-directed DNA methylation and Pol IV in Arabidopsis. Embo J 25 2828 2836
43. JiangDYangWHeYAmasinoRM 2007 Arabidopsis relatives of the human Lysine-Specific Demethylase 1 repress the expression of FWA and FLOWERING LOCUS C and thus promote the floral transition. Plant Cell 19 2975 2987
44. LippmanZMayBYordanCSingerTMartienssenR 2003 Distinct mechanisms determine transposon inheritance and methylation via small interfering RNA and histone modification. PLoS Biol 1 e67
45. ZhengBWangZLiSYuBLiuJY 2009 Intergenic transcription by RNA polymerase II coordinates Pol IV and Pol V in siRNA-directed transcriptional gene silencing in Arabidopsis. Genes Dev 23 2850 2860
46. ChanSWHendersonIRZhangXShahGChienJS 2006 RNAi, DRD1, and histone methylation actively target developmentally important non-CG DNA methylation in Arabidopsis. PLoS Genet 2 e83
47. ChanSWZilbermanDXieZJohansenLKCarringtonJC 2004 RNA silencing genes control de novo DNA methylation. Science 303 1336
48. CaoXJacobsenSE 2002 Role of the Arabidopsis DRM methyltransferases in de novo DNA methylation and gene silencing. Curr Biol 12 1138 1144
49. WuKZhangLZhouCYuCWChaikamV 2008 HDA6 is required for jasmonate response, senescence and flowering in Arabidopsis. J Exp Bot 59 225 234
50. Bracha-DroriKShichrurKKatzAOlivaMAngeloviciR 2004 Detection of protein-protein interactions in plants using bimolecular fluorescence complementation. Plant J 40 419 427
51. ToTKKimJMMatsuiAKuriharaYMorosawaT 2011 Arabidopsis HDA6 regulates locus-directed heterochromatin silencing in cooperation with MET1. PLoS Genet 7 e1002055
52. GaoZLiuHLDaxingerLPontesOHeX 2010 An RNA polymerase II- and AGO4-associated protein acts in RNA-directed DNA methylation. Nature 465 106 109
53. PazhouhandehMMolinierJBerrAGenschikP 2011 MSI4/FVE interacts with CUL4-DDB1 and a PRC2-like complex to control epigenetic regulation of flowering time in Arabidopsis. Proc Natl Acad Sci U S A 108 3430 3435
54. ZhangXClarenzOCokusSBernatavichuteYVPellegriniM 2007 Whole-genome analysis of histone H3 lysine 27 trimethylation in Arabidopsis. PLoS Biol 5 e129
55. SchubertDPrimavesiLBishoppARobertsGDoonanJ 2006 Silencing by plant Polycomb-group genes requires dispersed trimethylation of histone H3 at lysine 27. Embo J 25 4638 4649
56. JiangDWangYWangYHeY 2008 Repression of FLOWERING LOCUS C and FLOWERING LOCUS T by the Arabidopsis Polycomb repressive complex 2 components. PLoS ONE 3 e3404
57. FleischerTCYunUJAyerDE 2003 Identification and characterization of three new components of the mSin3A corepressor complex. Mol Cell Biol 23 3456 3467
58. ZhangYNgHHErdjument-BromageHTempstPBirdA 1999 Analysis of the NuRD subunits reveals a histone deacetylase core complex and a connection with DNA methylation. Genes Dev 13 1924 1935
59. HakimiMADongYLaneWSSpeicherDWShiekhattarR 2003 A candidate X-linked mental retardation gene is a component of a new family of histone deacetylase-containing complexes. J Biol Chem 278 7234 7239
60. ShiYLanFMatsonCMulliganPWhetstineJR 2004 Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 119 941 953
61. YuCWLiuXLuoMChenCLinX 2011 HISTONE DEACETYLASE6 interacts with FLOWERING LOCUS D and regulates flowering in Arabidopsis. Plant Physiol 156 173 184
62. JiangDGuXHeY 2009 Establishment of the winter-annual growth habit via FRIGIDA-mediated histone methylation at FLOWERING LOCUS C in Arabidopsis. Plant Cell 21 1733 1746
63. TanakaMKikuchiAKamadaH 2008 The Arabidopsis histone deacetylases HDA6 and HDA19 contribute to the repression of embryonic properties after germination. Plant Physiol 146 149 161
64. TianLWangJFongMPChenMCaoH 2003 Genetic control of developmental changes induced by disruption of Arabidopsis HISTONE DEACETYLASE 1 (AtHD1) expression. Genetics 165 399 409
65. MichaelsSAmasinoR 1999 FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 11 949 956
66. AlonsoJM 2003 Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653 657
67. EarleyKWHaagJRPontesOOpperKJuehneT 2006 Gateway-compatible vectors for plant functional genomics and proteomics. Plant J 45 616 629
68. CurtisMDGrossniklausU 2003 A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol 133 462 469
69. KarimiMDe MeyerBHilsonP 2005 Modular cloning in plant cells. Trends Plant Sci 10 103 105
70. GuXJiangDWangYBachmairAHeY 2009 Repression of the floral transition via histone H2B monoubiquitination. Plant J 57 522 533
71. JiangDKongNCGuXLiZHeY 2011 Arabidopsis COMPASS-like complexes mediate histone H3 lysine-4 trimethylation to control floral transition and plant development. PLoS Genet 7 e1001330
72. JohnsonLCaoXJacobsenS 2002 Interplay between two epigenetic marks: DNA methylation and histone H3 lysine 9 methylation. Curr Biol 12 1360 1367
73. SearleIHeYTurckFVincentCFornaraF 2006 The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev 20 898 912
Štítky
Genetika Reprodukčná medicínaČlánok vyšiel v časopise
PLOS Genetics
2011 Číslo 11
- Je „freeze-all“ pro všechny? Odborníci na fertilitu diskutovali na virtuálním summitu
- Gynekologové a odborníci na reprodukční medicínu se sejdou na prvním virtuálním summitu
Najčítanejšie v tomto čísle
- Evidence-Based Annotation of Gene Function in MR-1 Using Genome-Wide Fitness Profiling across 121 Conditions
- De Novo Origins of Human Genes
- Capture of MicroRNA–Bound mRNAs Identifies the Tumor Suppressor miR-34a as a Regulator of Growth Factor Signaling
- TRY-5 Is a Sperm-Activating Protease in Seminal Fluid