
Heritable Change Caused by Transient Transcription Errors
Transmission of cellular identity relies on the faithful transfer of information from the mother to the daughter cell. This process includes accurate replication of the DNA, but also the correct propagation of regulatory programs responsible for cellular identity. Errors in DNA replication (mutations) and protein conformation (prions) can trigger stable phenotypic changes and cause human disease, yet the ability of transient transcriptional errors to produce heritable phenotypic change (‘epimutations’) remains an open question. Here, we demonstrate that transcriptional errors made specifically in the mRNA encoding a transcription factor can promote heritable phenotypic change by reprogramming a transcriptional network, without altering DNA. We have harnessed the classical bistable switch in the lac operon, a memory-module, to capture the consequences of transient transcription errors in living Escherichia coli cells. We engineered an error-prone transcription sequence (A9 run) in the gene encoding the lac repressor and show that this ‘slippery’ sequence directly increases epigenetic switching, not mutation in the cell population. Therefore, one altered transcript within a multi-generational series of many error-free transcripts can cause long-term phenotypic consequences. Thus, like DNA mutations, transcriptional epimutations can instigate heritable changes that increase phenotypic diversity, which drives both evolution and disease.
Published in the journal:
Heritable Change Caused by Transient Transcription Errors. PLoS Genet 9(6): e32767. doi:10.1371/journal.pgen.1003595
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1003595
Summary
Transmission of cellular identity relies on the faithful transfer of information from the mother to the daughter cell. This process includes accurate replication of the DNA, but also the correct propagation of regulatory programs responsible for cellular identity. Errors in DNA replication (mutations) and protein conformation (prions) can trigger stable phenotypic changes and cause human disease, yet the ability of transient transcriptional errors to produce heritable phenotypic change (‘epimutations’) remains an open question. Here, we demonstrate that transcriptional errors made specifically in the mRNA encoding a transcription factor can promote heritable phenotypic change by reprogramming a transcriptional network, without altering DNA. We have harnessed the classical bistable switch in the lac operon, a memory-module, to capture the consequences of transient transcription errors in living Escherichia coli cells. We engineered an error-prone transcription sequence (A9 run) in the gene encoding the lac repressor and show that this ‘slippery’ sequence directly increases epigenetic switching, not mutation in the cell population. Therefore, one altered transcript within a multi-generational series of many error-free transcripts can cause long-term phenotypic consequences. Thus, like DNA mutations, transcriptional epimutations can instigate heritable changes that increase phenotypic diversity, which drives both evolution and disease.
Introduction
Stable phenotypic change is mostly associated with DNA alteration [1], the hardware of the cell, but rarely as the consequence of errors in the transmission of cellular genetic programs, the software of the cell [2], [3]. Transcription factors play a critical role in establishing cellular programs and heritable cellular identity [4], as elegantly shown by somatic cell nuclear transfer [5] and more recently reprogramming of differentiated cells into pluripotent cells [6]. Among cellular genetics programs, bistable gene networks play an important role in cellular differentiation and identity by allowing expression of multiple stable and heritable phenotypes from one single genome [7]. Examples of bistable systems include the E. coli lactose-operon-repressor system [8], the lambda bacteriophage lysis–lysogeny switch [9], the genetic toggle switch in bacteria [10] and human cells [11], phosphate response in yeast [12], cellular signal transduction pathways in Xenopus [13], HIV virus development [14], and the “restriction point”: the critical switch by which mammalian cells commit to proliferation and become independent of growth stimulation in cancer [15]. Recent studies on single cell genealogy analysis have revealed that heritable stochastic change can occur by dysregulation of bistable regulatory networks [16], [17]. This phenotypic switching has been associated with infrequent large bursts in transcription, generated by the stochastic dissociation of a transcription factor from its DNA regulatory site and resulting in a change of expression pattern [17].
Our previous studies suggested that transcription infidelity could contribute to stochastic heritable phenotypic change in a bistable gene network [18]. We showed that the removal of transcription fidelity factors in the cell, GreA and GreB (functional analogs of eukaryotic TFIIS), triggers heritable stochastic change [18]. Therefore, we proposed that an overall decrease in transcription fidelity affecting all nascent transcripts in the cell can increase stochastic switching in this system by altering the quality of the transcription factor involved in the bistable switch. However, these transcription fidelity factors also have other functions in transcription initiation and elongation [19]. In addition, a global decrease in transcription fidelity may indirectly trigger phenotypic switching by globally impacting the physiology of the cell, instead of directly altering the transcription factor mRNA [20], [21]. Due to the unstable nature of mRNA, the direct capture of the erroneous mRNA responsible for the phenotypic switch is not currently possible since by the time the cell exhibits the new phenotype, the initial erroneous mRNA will have been degraded. Hence, we have now developed a novel genetic approach to show that mRNA errors specifically in a transcription factor involved in a bistable switch can directly trigger heritable phenotypic change in a clonal cell lineage.
In E. coli, the lac operon has been shown to behave like a bistable switch and is considered a model system of gene regulation. The lac operon comprises a positive feedback loop that allows bistability under a specific concentration of inducer, thio-methylgalactoside (TMG), known as the maintenance concentration [8]. The lactose permease protein (encoded by the lacY gene) transports its own inducer, which in turn activates permease synthesis by derepressing its operon via inactivation of the lac repressor. Due to this autocatalytic positive feedback loop the lac operon exhibits two persistent and heritable expression states depending on the cellular history [8], [18], [22]. In the presence of the maintenance level of inducer, cells with permease will stay induced (ON) and cells without permease will stay uninduced (OFF) but will have a probability of switching ON. In their classic experiment, Novick and Weiner showed the persistence of the two heritable expression states for over 180 generations in a chemostat [8].
To directly test that transcription errors in the mRNA of a transcription regulator can promote phenotypic change, we engineered a transcriptional error-prone sequence into the lacI repressor gene, which dictates the fate of a heritable ON/OFF epigenetic switch in the lac operon. If our model that transcription errors cause epigenetic switching is correct, we predict that this engineered transcription error-prone sequence would lead to increased epigenetic ON-switching in the bistable lac system.
Results and Discussion
Engineering an Error-prone Sequence in the lacI Transcript
To directly look at the consequence of transcription errors in lacI mRNA, we elongated the native A3 sequence at the 5′ end of the chromosomal lacI gene to an A9 run or an A5GA3 control sequence (Figure 1A; Figure S4A) and thereby created a novel lac repressor that has two additional N-terminal appended Lys residues. Monotonous runs of adenines in DNA are hotspots for RNA polymerase slippage events during transcription in E. coli [23]–[26], Thermus thermophilus [27], yeast [25], [28] and human cells [29]–[31]. During transcription, the ∼8 bp hybrid between the nascent RNA chain and the DNA template maintains the proper register of the RNA transcript [32]. With a T9 run in the template (and an A9 run in the coding strand, as is found in our study), the growing chain with eight or nine A residues can dissociate from the template and realign out of frame, while still maintaining the required 8 bp RNA∶DNA hybrid. Therefore, the minimum length of the T run to promote transcriptional slippage is T9 [24], [27], [28]. In a wild-type gene, this transcriptional slippage results in the addition or deletion of a ribonucleotide in the A run in the transcript, resulting in a shift in the open reading frame and producing a burst of nonfunctional truncated protein.

Single cell analysis by flow cytometry shows that the GFP fluorescence histograms produced from OFF and ON populations of wild-type, A9 and A5GA3 repressor strains are identical demonstrating that the engineered Lys-Lys addition does not perturb repressor function (Figure 1B). The altered repressors still recognize and bind the lac operator, negatively regulate the lac operon and remain responsive to TMG, which binds to the structurally distinct C-terminal core domain and induces an allosteric transition in lac repressor so that it no longer binds to the lac operator. Moreover, all three repressor allele strains (wild-type, A9 and A5GA3) exhibit the same basal level of β-galactosidase activity in OFF populations indicating that the altered repressors bind as well as the wild-type repressor to the lac operator (Figure S4B). In addition, there is no difference in the spontaneous lacI+→lacI− mutation frequency for these two altered lacI alleles (Figure 1C; Text S1). Finally, the A9 and A5GA3 repressor/lac operon gene networks also exhibit bistability and hysteresis as has been shown for the native lac system (Figures 2, 3A,B; Figure S4C) [8], [18], [22]. To interrogate a larger number of cells, we used flow cytometry (Figure S1) and validated this method by showing that the frequencies of switching in diverse genetic backgrounds are similar to what we previously published (Figure S2). Thus, the discrimination afforded by flow cytometry analysis of GFP fluorescence between ON and OFF cells for the wild-type and variant lacI alleles is sensitive and sufficient to monitor stochastic switching.
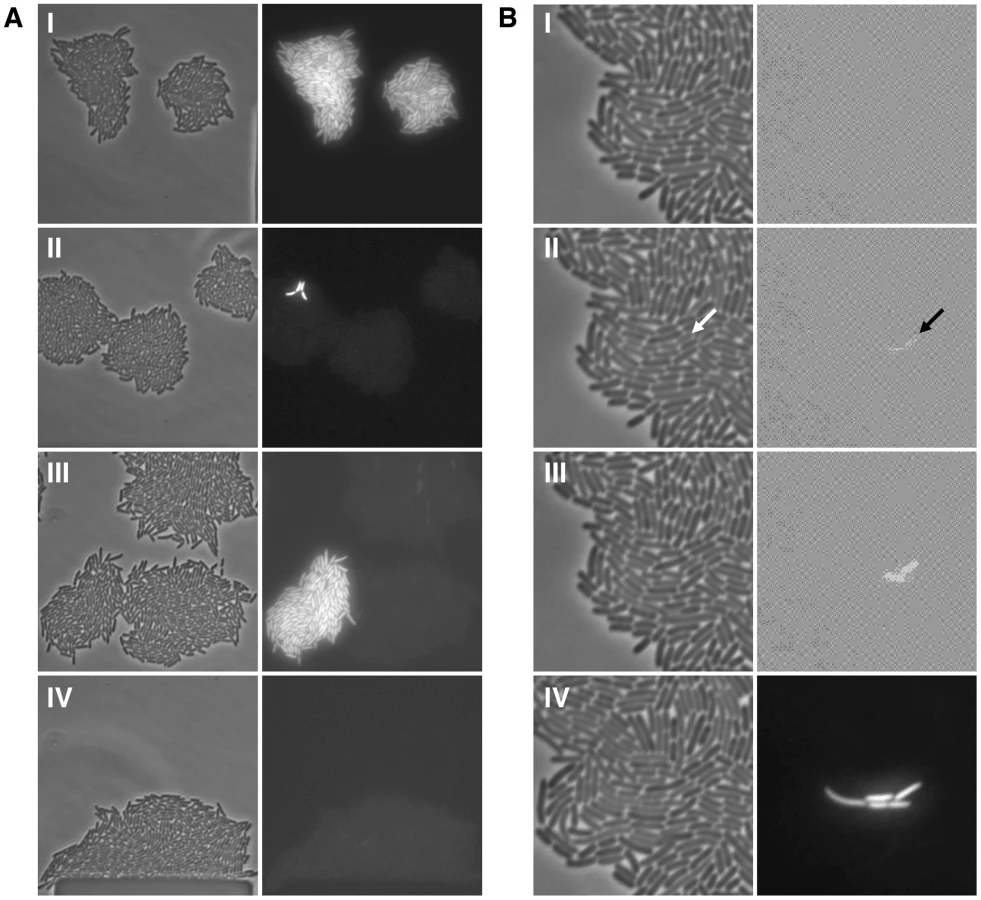

Transcriptional Slippage in the lacI Gene Increases Epigenetic Stochastic Switching
To determine the proportion of cells that are ON, we used the green fluorescent protein gene integrated within the lac operon (Figure 1A) [18]. During growth of OFF cells in a maintenance concentration of TMG, if a cell suffers a stochastic event leading to derepression of the lac operon, e.g. a transcription slippage error in lacI, the lac operon will be transiently derepressed triggering permease synthesis and activation of the autocatalytic positive-feedback loop, resulting in green fluorescent cells [18]. As a result, the OFF state will transition to the ON state and be heritably maintained in the following generations, mimicking lacI mutation in this system (i.e. transient stochastic events in information transfer can have heritable phenotypic consequences; Figure 2A,B) [8], [18]. We calculate the switch frequency as number of ON cells over the total number of cells interrogated, following the convention used in determining lacI− mutation frequencies [33]. The observed ON switch frequency is therefore dependent on both the number of switch events that have occurred and the number of generations after a discrete switch event has occurred, as in a classical fluctuation test; our experiments run for ∼28 bacterial generations (see Materials and Methods).
Between the two strains harbouring A9 and A5GA3 alleles, we can assess how slippery transcription sequences affect stochastic switching in the bistable lac system. We observe a 6 to 12-fold increase in switch frequency (Mann-Whitney Rank Sum Test, p<0.001) for the A9 construct compared with the A5GA3 control in a wild-type background at the maintenance concentration of TMG (Figure 3C,D; Figure S5). This A9 epigenetic switch frequency is fully 10,000 times greater than the genetic lacI− mutation frequency demonstrating that it is not mutation underlying the observed stochastic switching.
In addition, the observed increase in phenotypic switch frequency cannot be explained by problems in transcription initiation or early termination for the following reasons. First, native lacI mRNA, produced from a weak constitutive promoter, includes a 28 nucleotide (nt) untranslated leader sequence before the GTG initiation codon [34] and the A9 run (or the A5GA3 broken run) should not affect transcription initiation. Since the nascent transcript is 31 nt in length before the A9 run is first encountered, transcription of this sequence will be the processive synthesis of RNA as an RNA polymerase-DNA transcription complex and not as an RNA polymerase-promoter initial transcribing complex [35]. Second, aborted transcripts, if they occur from the lacI promoter, are also not relevant here; abortive events occur within 8–15 nt from the promoter, far away from the A9 (and the A5GA3) sequence. The absence of GreA function increases transcription abortion [35], however, we see no difference in transcript elongation in lacI monitored by a lacIZYA fusion in the absence of GreA, GreB or GreA,B function with these constructs whereas we see a significant increase in phenotypic switching in the absence of GreA,B (Figure 4). Third, we show that ±1 frameshifting in the coding sequence of lacI does not cause early transcription termination before downstream transcription into the lac operon (Figure S6). Together these experiments suggest that perturbations in transcription initiation and early termination are not involved in the increased OFF to ON switching frequency we observe. The net decrease in functional repressor in a cell, due to transcription error, may further require the dilution of ‘old’ wild-type repressors via partitioning during cell division to promote stochastic switching [36].

Transcription not Translation Errors Influence Stochastic Switching
To gain experimental evidence that transcription error and not translational frameshifting promotes stochastic switching in the A9 lacI allele, we introduced the A9 and A5GA3 lacI alleles into a ΔgreAB strain that contains deletions of the greA and greB genes that encode auxiliary fidelity factors that facilitate the proofreading of misincorporations that arise in nascent mRNAs during transcription [37]–[39]. We reasoned that if the switching is due to transcription slippage, removing transcription fidelity factors GreAB may promote more slippage and increase phenotypic switching. The increase in switch frequency in the presence of an error-prone sequence in the absence of RNA editing function is more than additive: the median switch frequency for the A9 allele in the ΔgreAB background (42.4%) is greater than the median switch frequency for the A9 allele (2.46%) and the ΔgreAB A5GA3 background (26.7%) combined (Figure 5A,B). When a slippery transcribed sequence is in a sloppy RNA transcription fidelity background, the switching frequency is significantly increased (Mann-Whitney Rank Sum Test, p<0.001), suggesting that misalignments due to transcriptional frameshifting are prevented by RNA editing of the Gre factors via transcription back-tracking/correction and support the role of transcription slippages in phenotypic change.

Furthermore, phenotypic switching has been associated with a large burst in permease synthesis [17]. An average gene transcript in E. coli will produce about 20–40 full-length polypeptides [40], [41]. Therefore, one transcription frameshift will produce 20–40 non-functional lac repressor sub-units (lac repressor is a tetramer), while one translational frameshift event will produce only one non-functional lac repressor sub-unit along with 20–40 functional lac repressor sub-units; a ratio of at least 20∶1 of functional over non-functional lac repressor sub-units will be the result. Therefore, the effects of a transcription error will be amplified nonlinearly over the original stochastic event generating tens of aberrant lac repressor polypeptides allowing a large burst of permease that is required for phenotypic change [17].
To measure the effect of translation frameshifting on lac operon induction, we created merodiploids that provide a 10-fold excess of wild-type transcripts over ±1 frameshift transcripts derived from the lacI A8 and A10 alleles we constructed on the chromosome (Table S2). We introduced an F′ factor with a wild-type lacI gene under control of the Iq up-promoter mutation, producing 10-fold more lacI transcription [42], into recA derivatives of our A8 and A10 frameshifted lacI strains (and into recA derivatives of our A9 and A5GA3 in-frame lacI strains; see Table S2). Therefore, the ratio of wild-type transcript over frameshifted transcript, at the level of transcription (10∶1), will be a very conservative approximation of the situation that would arise if during the translation of one A9 transcript, one translational frameshift event would occur since a ratio of at least 20∶1 of wild-type repressor sub-units over frameshifted sub-units would result. As shown in Figure 5C,D,E, a translation error on a pristine mRNA producing just one aberrant polypeptide amongst many wild-type polypeptides has no large effect on lac operon induction, demonstrating that more than one frameshifted repressor sub-unit is required to promote phenotypic switching.
Finally, efficient translation frameshifting is dependant on specific downstream sequence elements in E. coli [27] that are not obvious in our A9 lacI construct.
All together, these results show that insertion of a known transcription slippage sequence in the lacI transcript increases phenotypic switching in the bistable lac system due to transcription error in the mRNA and accumulation of a frameshifted and non-functional lac repressor leading to a transcription burst of lac permease.
Stochastic Errors in Information Transfer Have Heritable Phenotypic Consequences
DNA makes RNA makes protein; until now, errors in making two of the three elements in information transfer, DNA replication and protein folding, have been shown to modify cellular inheritance through mutation or prion conformational change [43] (Figure 6). Our results show that acute errors in mRNA, the transient element in information transfer, can also effect heritable change when they affect transcription factors involved in bistable gene networks. Transcription errors have been shown to have phenotypic consequences for the cell, but in the cases reported so far, chronic transcription errors can provide partial function or ‘leakiness’ giving an altered phenotype from a mutant or wild type gene [29]–[31], [44]–[46]. For example, a −1 frameshift mutation in the apolipoprotein B gene in a polyA run caused hypobetalipoproteinemia and transcription slippage at the polyA track restores the reading frame by insertion of an additional A and ameliorates the disease [29], [30]. In Alzheimer's patients, it was found that −2 frameshifts accumulated in amyloid precursor and ubiquitin B transcripts over time, which are thought to be important in nonfamilial early - and late-onset forms of Alzheimer's disease [47]. In contrast to these examples of chronic transcriptional errors producing a partial phenotype, we show that one acute transcription error on a poorly transcribed mRNA may promote heritable phenotypic change due to a change of connectivity in a transcription network. Although epimutation, a heritable change in gene expression that does not affect the actual base pair sequence of DNA, is usually associated with methylation patterns and epigenetic silencing of gene expression, the heritable stochastic switching due to transient transcription error we observe here, may also be included as epimutation.

RNA transcription errors are inevitable, ubiquitous and frequent (about 10,000 times more frequent than DNA replication errors) [48] and bistable gene networks sensitive to stochastic fluctuation in protein level are found in bacteria [17], yeast [49], fly [50], Xenopus [13], mammalian stem cells [51] and viruses such as HIV and lambda [52]. Furthermore, recent studies in E. coli [53], yeast [54] and human cells [55], [56] have shown that a majority of proteins involved in transcription networks are usually present at low abundance so it is easy to imagine that errors in one low abundant mRNA can have a drastic reduction in the protein concentration and trigger epigenetic change. So far, permanent activation or removal of transcription factors by genetic manipulation has been shown to promote a stable change of phenotype in a process known as transdifferentiation [57] and we speculate that transient disappearance of a transcription factor involved in a bistable switch may have the same phenotypic effect. Although here we focus on a slippery A9 sequence in the lacI gene, any kind of transcription error along the lacI transcript that would generate a non-functional lac repressor, may also initiate a stochastic switching event. Indeed, any transcribed DNA sequence that is problematic for RNA polymerase, due to a mono-, di-, tri - or some higher order nucleotide repeat or to hairpin or other secondary structures would produce a variety of mRNA species from the same gene. Therefore, transcription errors, epimutation, might be a general mechanism to create epigenetic heterogeneity in a clonal cell population and should be considered as one of the origins of phenotypic change that could lead to altered or aberrant cell behavior, impacting human health, including cancer if the dysregulation of key genetic bistable networks are altered by errors in transcription [58].
Materials and Methods
Bacterial Strains
All strains used in this study are derived from the wild-type sequenced E. coli MG1655 strain (Table S2). Manipulation of the MG1655 genome was accomplished by standard methodologies [33], [59]. To monitor the proportion of cells that are ON or OFF for lac operon expression, we have replaced the lacA gene in the wild-type E. coli MG1655 chromosome with a gfp cassette, so that when the lacZYA::gfp transcript is expressed, β-galactosidase, galactoside permease, and green fluorescent protein are produced from the lacZ, lacY, and gfp genes, respectively.
Growth Conditions and Media
To demonstrate hysteresis and bistability in lac operon expression in single cells, a bacterial culture grown in minimal A salts [33] plus MgSO4 (1 mM) with succinate (0.2%), was diluted 1∶5 in fresh medium with (ON culture) or without 1 mM TMG (OFF culture) and shaken at 37°C for 7 h. After this induction period, the two cultures were individually diluted and ∼200 cells were seeded to new tubes containing fresh medium that contained varying amounts of TMG, and shaken at 37°C for 42 h (∼28 generations). Flow cytometry was used to determine the percentage of cells that were induced for lac operon expression (ON cells).
To determine epigenetic and genetic switch frequencies, a bacterial culture grown in minimal succinate media, was diluted and ∼200 cells were seeded to new tubes containing fresh medium, with (epigenetic assay) or without (genetic assay) a maintenance level of TMG, and shaken at 37°C for 42 h. To determine genetic-mutation frequencies, dilutions of the subcultures were spread on selection plates (minimal A medium supplemented with 75 µg/ml phenyl-β-D-galactoside [Pgal] and 1.5% purified agar) and minimal A glucose (0.2%) plates and incubated for 2–3 days at 37°C. Pgal is a substrate of β-galactosidase and can act as a carbon source but does not induce lac operon expression, therefore only cells constitutively expressing β-galactosidase (lacI−) can form colonies [33].
Flow Cytometry
To determine the percentage of cells that were induced for lac operon expression (ON cells), 10 µl of culture was diluted into 300 µl filtered minimal A salts plus MgSO4 (1 mM) and subjected to flow cytometry analysis with GFP fluorescence measured in a BD FACSCanto II Flow Cytometer (Becton, Dickinson and Company, USA) with Diva acquisition software (Becton Dickinson) and FloJo analysis software (Tree Star, Inc. USA). To monitor fluorescent cells in a culture we used a narrow gating for forward and side scattering so that the most represented cell population was evaluated (Figure S1). For each independent culture 10,000 cells were interrogated.
Microscopy and Microfluidics
To follow the growth of single cells into microcolonies we used the CellASIC ONIX Microfluidic Platform (Millipore) including microfluidic perfusion system, microfluidic flow chamber for bacteria (BO4A plates) and FG software. Time-lapse microscopy was performed using a Zeiss HAL100 inverted fluorescence microscope. Fields were acquired at 100× magnification with an EM-CCD camera (Hammamatsu). Bright field and fluorescence (EGFP cube = Chroma, #41017; X-Cite120 fluorescence illuminator [EXFO Photonic Solutions]) images were acquired and image analysis was performed using AxioVision Rel. 4.6 (Zeiss). To maintain a constant 37°C environment throughout the experiment, the microscope was housed in an incubation system consisting of Incubator XL-S1 (PeCon) controlled by TempModule S and Heating Unit XL S (Zeiss).
Supporting Information
Zdroje
1. RandoOJ, VerstrepenKJ (2007) Timescales of genetic and epigenetic inheritance. Cell 128 : 655–668 doi:10.1016/j.cell.2007.01.023
2. DrummondDA, WilkeCO (2009) The evolutionary consequences of erroneous protein synthesis. Nat Rev Genet 10 : 715–724 doi:10.1038/nrg2662
3. SatoryD, GordonAJ, HallidayJA, HermanC (2011) Epigenetic switches: can infidelity govern fate in microbes? Curr Opin Microbiol 14 : 212–217 doi:10.1016/j.mib.2010.12.004
4. MonodJ, JacobF (1961) General conclusions: teleonomic mechanisms in cellular metabolism, growth, and differentiation. Cold Spring Harb Symp Quant Biol 26 : 389–401.
5. PasqueV, JullienJ, MiyamotoK, Halley-StottRP, GurdonJB (2011) Epigenetic factors influencing resistance to nuclear reprogramming. Trends Genet 27 : 516–525 doi:10.1016/j.tig.2011.08.002
6. TakahashiK, TanabeK, OhnukiM, NaritaM, IchisakaT, et al. (2007) Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131 : 861–872 doi:10.1016/j.cell.2007.11.019
7. DubnauD, LosickR (2006) Bistability in bacteria. Mol Microbiol 61 : 564–572 doi:10.1111/j.1365-2958.2006.05249.x
8. NovickA, WeinerM (1957) Enzyme induction as an all-or-none phenomenon. Proc Natl Acad Sci USA 43 : 553–566.
9. Ptashne M (2004) A genetic switch. New York: Cold Spring Harbor Laboratory Press. 164 p.
10. GardnerTS, CantorCR, CollinsJJ (2000) Construction of a genetic toggle switch in Escherichia coli. Nature 403 : 339–342 doi:10.1038/35002131
11. KalmarT, LimC, HaywardP, Muñoz-DescalzoS, NicholsJ, et al. (2009) Regulated fluctuations in Nanog expression mediate cell fate decisions in embryonic stem cells. PLoS Biol 7: e1000149 doi:10.1371/journal.pbio.1000149.g008
12. WykoffDD, RizviAH, RaserJM, MargolinB, O'SheaEK (2007) Positive feedback regulates switching of phosphate transporters in S. cerevisiae. Mol Cell 27 : 1005–1013 doi:10.1016/j.molcel.2007.07.022
13. FerrellJE, MachlederEM (1998) The biochemical basis of an all-or-none cell fate switch in Xenopus oocytes. Science 280 : 895–898.
14. WeinbergerLS, DarRD, SimpsonML (2008) Transient-mediated fate determination in a transcriptional circuit of HIV. Nat Genet 40 : 466–470 doi:10.1038/ng.116
15. YaoG, LeeTJ, MoriS, NevinsJR, YouL (2008) A bistable Rb–E2F switch underlies the restriction point. Nat Cell Biol 10 : 476–482 doi:10.1038/ncb1711
16. KaufmannBB, YangQ, MettetalJT, van OudenaardenA (2007) Heritable stochastic switching revealed by single-cell genealogy. PLoS Biol 5: e239 doi:10.1371/journal.pbio.0050239
17. ChoiPJ, CaiL, FriedaK, XieXS (2008) A stochastic single-molecule event triggers phenotype switching of a bacterial cell. Science 322 : 442–446 doi:10.1126/science.1161427
18. GordonAJE, HallidayJA, BlankschienMD, BurnsPA, YatagaiF, et al. (2009) Transcriptional infidelity promotes heritable phenotypic change in a bistable gene network. PLoS Biol 7: e44 doi:10.1371/journal.pbio.1000044.st002
19. StepanovaE, LeeJ, OzerovaM, SemenovaE, DatsenkoK, et al. (2007) Analysis of promoter targets for Escherichia coli transcription elongation factor GreA in vivo and in vitro. J Bacteriol 189 : 8772–8785 doi:10.1128/JB.00911-07
20. KaplanC (2009) Alternate explanation for observed epigenetic behavior? PLoS Biol 7: e44 Available: http://www.plosbiology.org/annotation/listThread.action?root=22157.
21. GordonAJE (2009) Pregnant pauses? PLoS Biol 7: e44 Available: http://www.plosbiology.org/annotation/listThread.action?root=9457.
22. OzbudakEM, ThattaiM, LimHN, ShraimanBI, Van OudenaardenA (2004) Multistability in the lactose utilization network of Escherichia coli. Nature 427 : 737–740 doi:10.1038/nature02298
23. ChamberlinM, BergP (1962) Deoxyribonucleic acid-directed synthesis of ribonucleic acid by an enzyme from Escherichia coli. Proc Natl Acad Sci USA 48 : 81.
24. WagnerLA, WeissRB, DriscollR, DunnDS, GestelandRF (1990) Transcriptional slippage occurs during elongation at runs of adenine or thymine in Escherichia coli. Nucleic Acids Research 18 : 3529–3535.
25. StrathernJN, JinDJ, CourtDL, KashlevM (2012) Isolation and characterization of transcription fidelity mutants. Biochim Biophys Acta 1819 : 694–699 doi:10.1016/j.bbagrm.2012.02.005
26. StrathernJ, MalagonF, IrvinJ, GotteD, ShaferB, et al. (2013) The fidelity of transcription: RPB1 (RPO21) mutations that increase transcriptional slippage in S. cerevisiae. J Biol Chem 288 : 2689–2699 doi:10.1074/jbc.M112.429506
27. LarsenB, WillsNM, NelsonC, AtkinsJF, GestelandRF (2000) Nonlinearity in genetic decoding: homologous DNA replicase genes use alternatives of transcriptional slippage or translational frameshifting. Proc Natl Acad Sci USA 97 : 1683–1688.
28. ZhouYN, LubkowskaL, HuiM, CourtC, ChenS, et al. (2013) Isolation and characterization of RNA Polymerase rpoB mutations that alter transcription slippage during elongation in Escherichia coli. J Biol Chem 288 : 2700–2710 doi:10.1074/jbc.M112.429464
29. LintonMF, PierottiV, YoungSG (1992) Reading-frame restoration with an apolipoprotein B gene frameshift mutation. Proc Natl Acad Sci USA 89 : 11431–11435.
30. LintonMF, RaabeM, PierottiV, YoungSG (1997) Reading-frame restoration by transcriptional slippage at long stretches of adenine residues in mammalian cells. J Biol Chem 272 : 14127–14132.
31. YoungM, InabaH, HoyerLW, HiguchiM, KazazianHH, et al. (1997) Partial correction of a severe molecular defect in hemophilia A, because of errors during expression of the factor VIII gene. Am J Hum Genet 60 : 565–573.
32. NudlerE, MustaevA, LukhtanovE, GoldfarbA (1997) The RNA-DNA hybrid maintains the register of transcription by preventing backtracking of RNA polymerase. Cell 89 : 33–41.
33. Miller JH (1992) A short course in bacterial genetics. New York: Cold Spring Harbor Laboratory Press. 456 p.
34. SteegeDA (1977) 5′-Terminal nucleotide sequence of Escherichia coli lactose repressor mRNA: features of translational initiation and reinitiation sites. Proc Natl Acad Sci USA 74 : 4163–4167.
35. GoldmanSR, EbrightRH, NickelsBE (2009) Direct detection of abortive RNA transcripts in vivo. Science 324 : 927–928 doi:10.1126/science.1169237
36. HuhD, PaulssonJ (2010) Non-genetic heterogeneity from stochastic partitioning at cell division. Nat Genet 43 : 95–100 doi:10.1038/ng.729
37. ErieDA, HajiseyedjavadiO, YoungMC, Hippel vonPH (1993) Multiple RNA polymerase conformations and GreA: control of the fidelity of transcription. Science 262 : 867–873.
38. ShaevitzJW, AbbondanzieriEA, LandickR, BlockSM (2003) Backtracking by single RNA polymerase molecules observed at near-base-pair resolution. Nature 426 : 684–687 doi:10.1038/nature02191
39. ZenkinN, YuzenkovaY, SeverinovK (2006) Transcript-assisted transcriptional proofreading. Science 313 : 518–520 doi:10.1126/science.1127422
40. KennellD, RiezmanH (1977) Transcription and translation initiation frequencies of the Escherichia coli lac operon. J Mol Biol 114 : 1–21.
41. CaiL, FriedmanN, XieXS (2006) Stochastic protein expression in individual cells at the single molecule level. Nature 440 : 358–362 doi:10.1038/nature04599
42. Müller-HillB, CrapoL, GilbertW (1968) Mutants that make more lac repressor. Proc Natl Acad Sci USA 59 : 1259–1264.
43. HalfmannR, JaroszDF, JonesSK, ChangA, LancasterAK, et al. (2012) Prions are a common mechanism for phenotypic inheritance in wild yeasts. Nature 482 : 363–368 doi:10.1038/nature10875
44. van LeeuwenF, van der BeekE, SegerM, BurbachP, IvellR (1989) Age-related development of a heterozygous phenotype in solitary neurons of the homozygous Brattleboro rat. Proc Natl Acad Sci USA 86 : 6417–6420.
45. BensonKF (2004) Paradoxical homozygous expression from heterozygotes and heterozygous expression from homozygotes as a consequence of transcriptional infidelity through a polyadenine tract in the AP3B1 gene responsible for canine cyclic neutropenia. Nucleic Acids Research 32 : 6327–6333 doi:10.1093/nar/gkh974
46. TaddeiF, HayakawaH, BoutonM, CirinesiA, MaticI, et al. (1997) Counteraction by MutT protein of transcriptional errors caused by oxidative damage. Science 278 : 128–130.
47. van LeeuwenFW (1998) Frameshift mutants of β amyloid precursor protein and Ubiquitin-B in Alzheimer's and Down patients. Science 279 : 242–247 doi:10.1126/science.279.5348.242
48. NinioJ (1991) Connections between translation, transcription and replication error-rates. Biochimie 73 : 1517–1523.
49. OzbudakEM, BecskeiA, van OudenaardenA (2005) A system of counteracting feedback loops regulates Cdc42p activity during spontaneous cell polarization. Developmental Cell 9 : 565–571 doi:10.1016/j.devcel.2005.08.014
50. WernetMF, MazzoniEO, ÇelikA, DuncanDM, DuncanI, et al. (2006) Stochastic spineless expression creates the retinal mosaic for colour vision. Nat Cell Biol 440 : 174–180 doi:10.1038/nature04615
51. MacArthurBen D, Ma'ayanA, LemischkaIR (2009) Systems biology of stem cell fate and cellular reprogramming. Nat Rev Mol Cell Biol 10 : 672–681 doi:10.1038/nrm2766
52. NachmanI, RamanathanS (2008) HIV-1 positive feedback and lytic fate. Nat Genet 40 : 382–383 doi:10.1038/ng0408-382
53. TaniguchiY, ChoiPJ, LiGW, ChenH, BabuM, et al. (2010) Quantifying E. coli proteome and transcriptome with single-molecule sensitivity in single cells. Science 329 : 533–538 doi:10.1126/science.1188308
54. HollandMJ (2002) Transcript abundance in yeast varies over six orders of magnitude. J Biol Chem 277 : 14363–14366 doi:10.1074/jbc.C200101200
55. VelculescuVE, MaddenSL, ZhangL, LashAE, YuJ, et al. (1999) Analysis of human transcriptomes. Nat Genet 23 : 387–388.
56. SigalA, MiloR, CohenA, Geva-ZatorskyN, KleinY, et al. (2006) Variability and memory of protein levels in human cells. Nature 444 : 643–646 doi:10.1038/nature05316
57. JoplingC, BoueS, BelmonteJCI (2011) Dedifferentiation, transdifferentiation and reprogramming: three routes to regeneration. Nat Rev Mol Cell Biol 12 : 79–89 doi:10.1038/nrm3043
58. BrulliardM, LorphelinD, CollignonO, LorphelinW, ThouvenotB, et al. (2007) Nonrandom variations in human cancer ESTs indicate that mRNA heterogeneity increases during carcinogenesis. Proc Natl Acad Sci USA 104 : 7522–7527 doi:10.1073/pnas.0611076104
59. DatsenkoKA, WannerBL (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97 : 6640–6645 doi:10.1073/pnas.120163297
60. OehlerS, EismannER, KrämerH, Müller-HillB (1990) The three operators of the lac operon cooperate in repression. EMBO J 9 : 973–979.
61. PlattT, WeberK, GanemD, MillerJH (1972) Translational restarts: AUG reinitiation of a lac repressor fragment. Proc Natl Acad Sci USA 69 : 897–901.
62. KleinaLG, MillerJH (1990) Genetic studies of the lac repressor. XIII. Extensive amino acid replacements generated by the use of natural and synthetic nonsense suppressors. J Mol Biol 212 : 295–318 doi:10.1016/0022-2836(90)90126-7
63. ClarkeND, BeamerLJ, GoldbergHR, BerkowerC, PaboCO (1991) The DNA binding arm of lambda repressor: critical contacts from a flexible region. Science 254 : 267–270.
64. SellittiMA, PavcoPA, SteegeDA (1987) lac repressor blocks in vivo transcription of lac control region DNA. Proc Natl Acad Sci USA 84 : 3199–3203.
65. AboT, InadaT, OgawaK, AibaH (2000) SsrA-mediated tagging and proteolysis of LacI and its role in the regulation of lac operon. EMBO J 19 : 3762–3769.
Štítky
Genetika Reprodukčná medicínaČlánok vyšiel v časopise
PLOS Genetics
2013 Číslo 6
- Gynekologové a odborníci na reprodukční medicínu se sejdou na prvním virtuálním summitu
- Je „freeze-all“ pro všechny? Odborníci na fertilitu diskutovali na virtuálním summitu
Najčítanejšie v tomto čísle
- BMS1 Is Mutated in Aplasia Cutis Congenita
- Sex-stratified Genome-wide Association Studies Including 270,000 Individuals Show Sexual Dimorphism in Genetic Loci for Anthropometric Traits
- Distinctive Expansion of Potential Virulence Genes in the Genome of the Oomycete Fish Pathogen
- Distinct Neuroblastoma-associated Alterations of Impair Sympathetic Neuronal Differentiation in Zebrafish Models