
Základy kognitivní, afektivní a sociální neurovědy: XVIII. Interocepce a emoční pozadí
Basics of social cognitive and affective neuroscience. XVIII. Interoception and emotional background
Damasio proposes that emotions are part of multi-tiered, hierarchical and evolutionary set neural mechanisms aimed at maintaining organism homeostasis. Part of this mechanism and so-called emotional background is Craig’s interoception, the sense of the physiological condition of the body. Primates have a distinct cortical image of homeostatic afferent activity in the posterior insula. In humans there exists metarepresentation of the primary interoceptive activity in the anterior insula and other parts of the brain. Interoceptive awareness drives increased rational decision-making and enhances neural activity during empathy.
Key words:
emotional background, interoception, interoceptive system, insula
Autori:
Koukolík; F
Pôsobisko autorov:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Thomayerova nemocnice, Praha
; Oddělení patologie a molekulární medicíny
Vyšlo v časopise:
Prakt. Lék. 2012; 92(5): 251-256
Kategória:
Editorial
Súhrn
Damasio předpokládá, že emoce jsou částí mnohovrstevného, hierarchického, evolučně daného neuronálního mechanismu, jehož smysl je uchování homeostázy organismu. Součástí tohoto mechanismu a tzv. emočního pozadí je Craigova interocepce, systém rozlišující fysiologický stav organismu. Aktivitu homeostatických aferent, která reflektuje fysiologický stav tkání a orgánů, reprezentují primáti v zadní insulární kůře. U lidí se prokazuje metareprezentace primární interoceptivní aktivity v přední insulární kůře a dalších částech mozku. Interoceptivní „vědomí“ ovlivňuje ekonomickou racionalitu a zvyšuje aktivitu insulární kůry v průběhu empatického procesu.
Klíčová slova:
emoční pozadí, interocepce, interoceptivní systém, insula
Úvod
Pravděpodobně díky antické, racionalistické a osvícenské myšlenkové tradici se lidé, alespoň v západním kulturním okruhu, považují za racionální bytosti. Tato a následující stati zabývající se současným poznáním stavby a činnosti „emočního mozku“ se pokusí doložit, že jsme v podstatně větší míře bytosti vysoce citové a myslící spíše magicky, a popsat důvody.
Definice a rozlišování emocí a pocitů jsou pojmová džungle (box 1). Pojem emoce se často užívá i pro pojem pocity. Problematické je vymezení pojmu afekt. Zajímavý výzkumný směr studia emocí se zračí v souboru prací uveřejňovaných Antoniem Damasiem a jeho spolupracovníky z Iowské univerzity přibližně od 90. let minulého století. Klasifikace emocí a pocitů uveřejňovaná touto školou je samozřejmě předmětem odborné debaty, je však výhodná didakticky. Damasio do jisté míry emoce od pocitů odděluje s tím, že emoce pocitům předcházejí (10–14).

Damasiova pracovní definice emocí říká: „emoce, například štěstí, smutek, znepokojení či pýcha, uspořádaný soubor chemických a neuronálních odpovědí mozku na emočně kompetentní podnět – například na objekt nebo situaci. Zpracování podnětu může, ale nemusí být vědomé, neboť emoční odpovědi jsou automatické. Emoční odpovědi jsou ty druhy odpovědí mozku, které evoluce připravila jako odpovědi na některé typy objektů a událostí společně s jistým rejstříkem akcí. Mozek nakonec asociuje další objekty a události, které jsou součástí individuální zkušenosti, s oněmi objekty a událostmi, které jsou příčinou emocí z vrozených důvodů. Tím vzniká další množina emočně kompetentních podnětů.“
Damasiova pracovní definice pocitů: „Pocity jsou mentální reprezentace fyziologických změn charakterizujících emoce. Na rozdíl od emocí, které jsou vědecky zjevné, jsou pocity skutečně jevem soukromým, byť nikoli subjektivnějším než jsou jakékoli jiné aspekty vědomí, jejichž příkladem je mé plánování této věty nebo mentální řešení aritmetického problému. Pocity jsou vědecky analyzovatelné stejně jako jakékoli jiné kognitivní jevy za předpokladu užití přiměřené metody. A nadto: protože jsou pocity přímým důsledkem emocí, vysvětlení neurobiologie emocí otevírá cestu k vysvětlení neurobiologie pocitů. Jestliže emoce zaručují okamžitou odpověď na jisté výzvy a příležitosti, jimž organismus čelí, pocit těchto emocí organismu oznamuje vědomý poplach. Pocity zesilují vliv dané situace, zvyšují míru učení, zvyšují pravděpodobnost, že bude možné podobnou situaci předvídat.“
Stať
Damasio vytvořil hierarchický model emocí a pocitů: „vlastní emoce“ (emotions-proper, mezi něž řadí emoční pozadí, dále primární neboli základní a sociální emoce) považuje Damasio za vysokou úroveň automatické homeostázy.
Jednotlivé úrovně automatické homeostázy jsou:
- základní úroveň, která odpovídá metabolické regulaci, základním reflexům, imunitní odpovědi;
- na druhé úrovni jsou chování spjatá s bolestí a slastí. Damasio upozorňuje, že zkušenost bolesti nebo slasti není příčinou chování, která bolesti nebo slasti odpovídají, a není pro tyto druhy chování nutná. Druhy chování, které lze označit v tomto pojetí jako emoční, lze stopovat u vývojově nižších živočichů, například u octomilek, aniž bychom předpokládali, že mají pocity;
- na třetí úrovni jsou instinktivní potřeby (drives) a motivace, hlavními příklady jsou hlad, žízeň, zvědavost a zkoumání, hra a sexuální chování;
- na čtvrté úrovni se objevují vlastní emoce. Kromě takzvaného emočního pozadí se mezi ně řadí základní (primary, basic) emoce. Součástí této skupiny je široká, rozličným způsobem tříděná množina sociálních emocí;
- nejvyšší úrovní jsou pocity (feelings) považované za mentální vyjádření všech „nižších“ úrovní homeostatické regulace – emoce vstupují do vědomí a sebeuvědomování v podobě pocitů (14).
Tato stať se věnuje klíčové složce emočního pozadí, interocepci. Její chápání se v posledních letech zásadně změnilo. Emoční pozadí (background emotions) lze chápat jako „kombinaci jednodušších regulačních reakcí, například základních homeostatických procesů, třeba metabolických adaptací, ale i chování spjatých s bolestí a slastí, stejně jako výsledků vyhodnocování proměn zevního prostředí zvládaných jinými emocemi, potřebami, případně i intelektuální rozvahou. Proměny emočního pozadí lze považovat za obtížně předvídatelné, vlivy, které jsou jejich příčinou i proměny samotné, jsou často protikladné“ (14).
Na své emoční pozadí se tážeme, jakmile se sami sebe zeptáme „jak se cítím?“ a vypovídáme o něm „cítím se dobře, cítím se špatně“.
Němečtí fyziologové z první poloviny 19. století chápali informace, které vnímáme ze svých těl a označujeme například pojmy teplota, bolest, svědění, lechtání, svalové napětí nebo ochablost, hlad, žízeň, za Gemeingefühl, „společný pocit“. Odlišovali je od smyslových informací zpracovávaných běžnými pěti smysly. Sherrington pro tento pojem užil formulaci „material me“: všechny tyto pocity jsou základem pocit vlastního jáství (5). Později však Sherrington rozdělil smysly na modality: teloreceptivní (zrak, sluch), proprioceptivní (poloha končetin), exteroceptivní (dotyk), chemoreceptivní (pachy, vůně a chutě) a interoceptivní (viscerální). Pocit teploty a bolest považoval za aspekty dotyku. Současný výzkum funkční anatomie spinothalamokortikálního systému vycházejícího z Rexedovy lamina I páteřní míchy dokazuje, že je nutné interocepci chápat jako fyziologickou funkci týkající se celého těla, nikoli jen vnitřních orgánů (4–6).
Interocepce je homeostatický aferentní systém přenášející signály ze všech tělesných tkání a orgánů prostřednictvím tenkých primárních aferent promítajících prostřednictvím neuronů lamina I do autonomních a homeostatických center míšních a kmenových. Společně s aferentními signály zprostředkovanými neurony nucleus tractus solitarius tvoří u primátů přímou thalamokortikální reprezentaci stavu těla (5, 23; obr. 1). Tato interoceptivní reprezentace je znovu reprezentována v rostrálních oblastech inzulární kůry pravé hemisféry, což vytváří – možná unikátně u člověka – základ pro subjektivní vyhodnocení vlastního stavu, onoho: „jak se cítím“ (4, 8, 9).
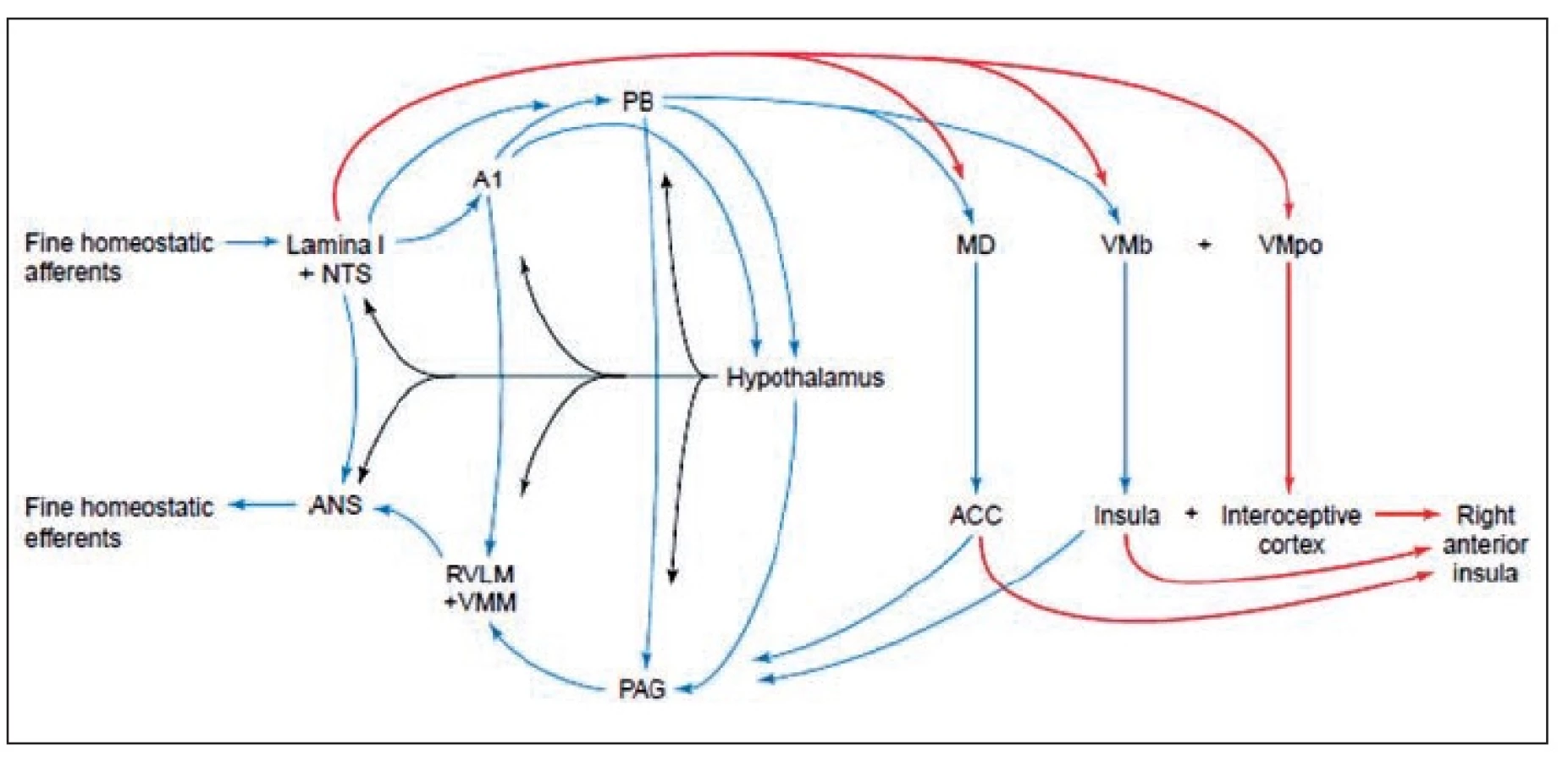
V porovnání s rozšířeným a vžitým učebnicovým Sherringtonovým pojetím jde o radikální názorovou změnu: bolest, teplota jsou stejně jako „pocity“ z vnitřních orgánů chápány jako homeostatická interocepce, nikoli exterocepce. Klíčovou skutečností odlišující bolest, teplotu a další tělesné pocity je jejich sepětí s emocemi. Příkladem může být pocit teploty reflektující stav tělesného povrchu i vnitřních orgánů, který s sebou nese neoddělitelný afekt závisející na stavu těla. Chladná voda s touž teplotou může být v letním vedru příjemná, jakmile jsme promrzlí na kost velmi nepříjemná. Rozlišování teploty proto v současnosti přestává být považováno za submodalitu dotykového čití. Craig et al. (5) pomocí PET prokázal, že stupňovitě chladnější podněty aktivují dorzální okraj kontralaterální střední a zadní inzuly. To odpovídá termoreceptivně a nociceptivní specifické spinothalamokortikální dráze vystupující u opic z Rexedovy lamina I spinální míchy. Dorzální okraj střední a zadní inzuly je proto považován za exteroceptivní oblast limbické kůry. Poškození této oblasti může být příčinou syndromu centrální bolesti. Ten může být důsledkem ztráty normální inhibice bolesti chladem. Subjektivní míra intenzity tepelného podnětu dobře odpovídala aktivaci stejnostranné (v tomto případě pravostranné) přední inzulární a orbitofrontální kůry.
Primární aferentní vlákna A-δ a C a Rexedova lamina I šedé hmoty spinální míchy
Primární aferentní vlákna A-δ a C (první jsou myelinizovaná, vedou tedy rychleji, než druhá, nemyelinizovaná), která inervují všechny tělesné tkáně, přinášejí homeostatické informace, tedy daleko víc než jen „pocit bolesti a tepla“. Dopravují informace o mechanickém stresu, o změnách lokálního metabolismu, například o pH, hypoxii, hyperkapnii, hypoglykémii, hypoosmolalitě, místní koncentraci laktátu, dále o buněčné ruptuře, průniku parazitů kůží, o aktivaci žírných buněk, o imunitní a hormonální aktivitě.
A-δ vlákna jsou slabě myelinizovaná, vedou středně rychle, C vlákna myelinizovaná nejsou, vodí pomalu. Mnohá kožní vlákna C jsou nadto selektivně citlivá na pomalé, slabé mechanické podněty probouzející pocit senzuálního („limbického“) dotyku. Monosynaptický vstup tenkých primárních aferentních vláken A-δ a C, která inervují všechny tělesné tkáně (uvádí se jedno zakončení v krychličce s hranou 15 mikrometrů), oslovuje neurony Rexedova laminy I. Ta je v nejpovrchnější neuronální zóně zadních rohů spinální míchy a trigeminálního vstupu (box 2).

Vlákna vystupující z neuronů lamina I tvoří přibližně polovinu spinálních vstupů do mozkového kmene a thalamu. Neurony lamina I, jimž se říká označené (labelled lines), to znamená anatomicky a fyziologicky vymezené neurony výlučně sloužící specifickému počitku, přenášejí informace, které interpretujeme jako teplotu a bolest.
Vzestupná vlákna neuronů lamina I vydávají v thorakolumbální míše kolaterály k neuronům sloupce sympatiku. V prodloužené míše pak promítají k neuronům jejích katecholoaminergních systémů, parabrachiálního jádra, šedi kolem mokovodu. Oblast je mohutně propojena s hypothalamem, amygdalou a thalamem. Ze dvou thalamických projekčních jader směřují vlákna do dorzální insulární kůry.
Dorzální inzulární kůra tedy u primátů reprezentuje aktivitu tenkých aferentních vláken odpovídající fyziologickému stavu celého těla. V rámci této aktivity jsou reprezentovány i vymezené pocity nazývané teplotou, bolestí, svěděním, svalovými a viscerálními pocity, včetně senzuálních dotyků (obr. 2).
Vyšetření PET ukazuje aktivaci pravostranné inzulární kůry v souvislosti s rozmanitými subjektivními pocity, jako jsou pocit hněvu vyvolaný vzpomínkami, pocit chladu odpovídající aplikaci chladného podnětu na ruku, prohlubující se výraz hnusu ve zrakově sledovaných tvářích, míra důvěryhodnosti tváří i sexuální nabuzení (4–6).
Reprezentace (funkční mapy) dorzální inzulární kůry jsou znovu reprezentovány v přední inzulární kůře stejnostranně a prostřednictvím kalosálních vláken i na straně opačné. Tato re-reprezentace je pak předávána do pravostranné orbitofrontální kůry. Stává se tím funkčním podkladem pocitů, který odpovídá Damasiově teorii. Je však nutno mít na mysli, že společně s přední inzulární kůrou se ve všech funkčních zobrazovacích studiích zkoumajících emoce současně aktivuje přední cingulární kůra, amygdala a ventrální striatum (16, 25, 2,1).
Critchley et al. (5) požádal zdravé dobrovolníky, aby věnovali pozornost buď frekvenci a rytmu vlastní srdeční akce, nebo tónu, který uslyší ve sluchátkách. Jakmile lidé věnovali pozornost vlastní srdeční akci, rostla aktivita mozkové kůry v inzule, somatomotorické a cingulární kůře. Míra aktivity pravostranné inzulární a operkulární kůry odpovídala míře přesnosti, se kterou vyšetřovaní lidé určovali vlastní srdeční rytmus. Objem šedé hmoty v této oblasti odpovídal míře subjektivního hodnocení a přesnosti, s níž si lidé byli vědomi akce vlastního srdce. S mírou přesnosti interoceptivního vnímání byly v korelaci indikátory záporných emočních zkušeností. Z toho plyne potvrzení Damasiovy predikce: aktivita přední části pravostranné inzulární kůry odpovídá viscerální reprezentaci přístupné vědomí. Tím se stává podkladem subjektivních pocitů.
Pollatosová et al. (26) při analogickém experimentu rozdělila zdravé dobrovolníky na skupinu, která vnímá vlastní srdeční akci dobře a skupinu, která ji dobře nevnímá. Oběma skupinám pak byly předvedeny příjemné, nepříjemné a neutrální obrázky z mezinárodního systému obrázků ovlivňujících afekt.
V průběhu prezentace byla snímána elektrokardiograficky srdeční akce, kromě toho elektroencefalogram. Po prezentaci každého obrázku vyšetřovaní lidé hodnotili v devítistupňové škále jeho emoční valenci a míru nabuzení. Lidé, kteří dobře rozlišovali vlastní srdeční akci, prožívali emočně působící obrázky v daleko větší míře, než lidé, kteří vlastní srdeční akci rozlišovali špatně. Valenci obrázků rozlišovaly obě skupiny stejně.
Mezi oběma skupinami byl zjištěn významný rozdíl elektroencefalografické aktivity. Amplituda vlny P300 lidí, kteří rozlišovali svou srdeční akci dobře, byla významně vyšší než u opačné skupiny. Mezi oběma skupinami byly rovněž zjištěny signifikantní rozdíly latence pomalé složky záznamu.
Weiss et al. (29) studoval odpověď mozku na selektivní stimulaci A-δ a C vláken funkční magnetickou resonancí. Na stimulaci C vláken odpovídala v porovnání se stimulací A-δ vláken kůra pravého frontálního opercula a přední insuly. Je tedy možné, že je funkce těchto dvou systémů odlišná, předpokládá se, že C vlákna vedou signály s větším emočním významem.
Čistá autonomní insuficience jako model
Podíl uvedených korových oblastí na rozlišování emocí dokládá i vyšetření nemocných s čistou autonomní insuficiencí (PAF, pure autonomic failure), což je onemocnění projevující se ve středním věku. Postihuje výlučně periferní vegetativní systém. V jeho gangliích se histologicky najdou Lewyho tělíska. Příčina není známa. Onemocnění se odlišuje od neurodegenerativních chorob centrálního nervového systému, které jsou rovněž doprovázeny insuficiencí vegetativního systému, například od multisystémové atrofie nebo Parkinsonovy nemoci. Očekávaná délka dožití je u pacientů s PAF stejná jako u kontrolní populace.
Klíčovým příznakem PAF je ortostatická hypotenze. V průběhu namáhavého cvičení nebo namáhavější mentální aktivity se u těchto pacientů nezrychluje tep a nestoupá krevní tlak, nestoupá hladina cirkulujících katecholaminů, na základě emočních podnětů nebo podnětů zatěžujících orientace se nemění elektrická kožní odpověď, je zmenšen pupilární reflex.
Critchley et al. (7) zatížil 9 pacientů s PAF a 8 zdravých lidí namáhavější a málo namáhavou tělesnou zátěží, dále namáhavějším a málo namáhavým řešením aritmetických problémů. Teoretický model předpovídá reprezentaci autoregulace nejprve na úrovni dorzální oblasti Varolova mostu (reprezentace prvního řádu), dále reprezentaci v oblasti inzuly a somatosenzorické, cingulární, případně i parietální kůry (reprezentace druhého řádu; 7).
Výsledky experimentu doložily teoretický předpoklad. Pacienti stižení PAF jsou poškozováni periferní autonomní denervací. Tenká vlákna jim tedy z periferie nepřinášejí informace. Centrální eferentní vegetativní kontrolu však mají uchovanou. Mají tedy „odpojenou“ zpětnou vazbu. Podle očekávání jim namáhavé cvičení ani namáhavá mentální zátěž nezvýšily krevní tlak ani frekvenci srdeční akce. Bez ohledu na druh úlohy se v porovnání se zdravými jedinci u těchto pacientů zvyšovala aktivita v oblasti dorzálního pontu, v místech, kde jsou jádra odpovídající za reprezentaci a kontrolu vegetativních odpovědí, zejména parabrachiální jádro a neurony šedi kolem mokovodu. Vyšší aktivita, opět bez ohledu na typ úlohy, byla v porovnání se zdravými lidmi zjištěna i v oblasti přední cingulární kůry, inzuly, thalamu a colicullus superior a somatosenzorické kůry. Naproti tomu nižší aktivita byla v porovnání se zdravými lidmi zjištěna v oblasti zadní cingulární a mediální parietální kůry. Pacienti s PAF v porovnání se zdravými lidmi udávají odchylky emocí vysokým skóre ve výrocích typu: „Již nejsem s to pociťovat smutek“, „Ztratil jsem schopnost pociťovat emoce“, „Cítím se všem lidem vzdálený“, „Mám pocit, že své emoce již nekontroluji“.
Insulární kůra má další funkce
Je prospěšné mít na vědomí, že se insula kromě homeostatických informací podílí na zpracovávání jazyka a motorické kontroly. Mutschler et al. (24) v metaanalytické studii šetřil topografické uspořádání insulární kůry a její vztah k amygdale, která je s insulární kůrou mohutně propojená. Sluchové a jazykové úlohy aktivovaly dorzální část přední insulární kůry. Motorické úlohy cílené na horní a dolní končetinu aktivovaly zadní část přední insulární kůry v blízkosti sulcus centralis insulae. Jestliže nějaký druh zátěže aktivoval amygdalu, aktivovala se současně ventrální část přední insulární kůry, která současně odpovídala na fysiologické podněty z periferie. Z toho plyne, že se dorzální část přední insulární kůry podílí na integraci sluchových a motorických informací, zatímco její část ventrální ovlivňuje fysiologické změny v průběhu rozličných emočních stavů.
Cauda et al. (3) v nové studii dokazuje, že je insula uzlem dvou komplementárních sítí, jedné ve spodní (ventrální) a přední části, druhé v horní (dorzální) a zadní části. Přední insula je propojena se střední a dolní spánkovou kůrou, dále s přední cingulární kůrou. Je tedy součástí limbického systému a podílí se na zpracovávání emocí. Střední, horní a zadní insulární kůra je propojena s premotorickou, senzoricko-motorickou, doplňkovou motorickou a střední a zadní cingulární kůrou, takže se podílí na integraci senzoricko-motorických funkcí. Kromě toho byla prokázána částečná lateralizace: například přední a dolní insulární kůra je propojena s cingulární kůrou mohutněji na straně pravé, na straně levé je naproti tomu mohutněji propojena s kůrou frontální. Zadní a horní insula je mohutně propojena se spánkovou a týlní kůrou vpravo. Lateralizace propojení odpovídá nálezům u primátů a má zatím nejasný funkční význam.
Craig (6) v přehledné studii dokládá:
- Interoceptivní podněty aktivující přední insulární kůru jsou žízeň, nedostatek vzduchu, dyspnoe, Valsalvův manévr, jemný, hladivý tzv. senzuální dotek, svědění, stimulace penisu, sexuální nabuzení, chlad, teplo, cvičení, sledování vlastního srdečního rytmu, ochutnávka vína u sommeliérů, roztažení močového měchýře, konečníku a jícnu. Přitom schopnost uvědomovat si vlastní srdeční rytmus odpovídá schopnosti rozlišovat vlastní emoce. Významným interoceptivním podnětem je bolest. Aktivita insulární kůry je součástí centrálního neuronálního mapování bolesti. Tepelný bolestivý podnět aplikovaný na hřbet ruky aktivuje kontralaterální zadní horní insulární kůru, střední insulární kůru vpravo víc než vlevo a jestliže se subjekt na podnět soustředí, aktivuje nadto přední insulární kůru, vpravo podstatně víc než vlevo. Tato část insulární kůry je tudíž součástí mapy sebeuvědomování. Jestliže lidé sledují bolestivé podněty, jimž jsou vystaveni bližní, které mají rádi, aktivuje se přední insulární kůra oboustranně. Tato korová oblast je tedy i součástí mapy emoční empatie.
- Kromě toho insulární kůru – jako ve všech uvedených případech jde vždy o součást neuronální sítě velkého rozsahu – aktivuje vědomí vlastního pohybu, sebeuvědomování, vokalizace a hudba, složitější emoce, příkladem je prožívání mateřské i romantické lásky, averze, pocit nespravedlivého rozdělování, sociální vyřazení, nerovnost, nejistota, důvěra, pocit prožitku krásy ze sochy, pocit prožitku jednoty s Bohem a halucinogenní stav navozený ayahuascou, což je odvar z „liány mrtvých“, obsahující směs psychoaktivně účinných látek obohacenou dalšími rostlinnými extrakty. Aktivaci insulární kůry způsobí vědomí rizika, anticipace a nejistota, zrakové a sluchové vědomí právě probíhajícího okamžiku, tedy rozlišování času, úlohy zatěžující pozornost, percepční rozhodování, kontrola poznávacích funkcí a jejich sledování.
I tyto experimenty dokládají obecně platící pravidlo: funkční systémy mozku je lépe chápat jako konektomy, než jako vymezené moduly. Stejné uzly mohou zprostředkovávat různé funkce a naopak: různé uzly mohou zprostředkovávat stejné funkce (19).
Interocepce a hra Konec smlouvání
Konec smlouvání je jednoduchá ekonomická hra pro dva hráče A, B. Hráč A má k dispozici nějakou finanční sumu. Hráč B její výši zná. Hráč A se rozhodne, jak ji rozdělí mezi sebe a hráče B. Za sociální normu se přitom považuje rozdělení stejným dílem. Hráč B buď nabízený podíl přijme, nebo odmítne. Jestliže přijme, je suma rozdělena způsobem, o kterém rozhodl A. Jestliže B odmítne, nedostane nikdo nic. Ekonomicky racionální jedinec by přijal jakkoli nízký podíl – lepší něco, než nic. Lidé se však tímto způsobem nechovají. Testování modelu v nejrůznějších kulturních okruzích s velmi rozmanitými částkami doložilo: čím víc se nabízený podíl blíží čtvrtině, tím roste pravděpodobnost odmítnutí. Lidé trestají chamtivého jedince za cenu vlastní ztráty. Teorie mluví o altruistickém trestání interpretovaném jako nástroj vynucující zachování sociální normy a udržení kooperace.
Nespravedlivé nabídky v ekonomických hrách tohoto typu ve zvýšené míře aktivují kůru přední insuly. Podobným způsobem ji aktivuje jak prožitek fyzického, tak morálního hnusu. Z toho plyne, že odmítnutí nefér nabídky je podmíněno emočně. Tím pádem by odmítání nefér nabídek mohlo být u lidí, dobře kontrolujících své emoce, odlišné. Mezi tyto lidi patří jedinci s meditační zkušeností.
Meditace má asi 2500 let dlouhou tradici. Lze ji chápat jako soubor postupů směřujících kromě jiného k emoční regulaci. Smyslem meditace je, opět kromě jiného, emoční vyrovnanost (21). Dlouhodobé meditování mění stavbu a funkci mozku (20). V průběhu meditačního stavu roste aktivita kůry střední a zadní insuly (22). Již krátký meditační výcvik dokáže „odpojovat“ aktivitu insuly od aktivity dalších korových oblastí, které se podílejí na sociálním rozhodování, například mediální prefrontální kůry (27). Lidé s delší meditační zkušeností by tedy měli na nespravedlivé nabídky ve hře Konec smlouvání odpovídat odlišně než lidé bez takové zkušenosti.
Kirk et al (17) požádal 26 lidí s meditační zkušeností dlouhou 6 měsíců – 24 let, muže a ženy o účast v experimentu, v jehož průběhu je zatížili 45 koly anonymní hry Konec smlouvání. Kontrolní skupinu tvořilo 40 lidí, opět žen i mužů, bez zkušenosti s meditací. Lidé s meditační zkušenosti přijali nefér nabídky přibližně v polovině případů, lidé, kteří tuto zkušenost neměli, přibližně ve čtvrtině případů. Nefér nabídka aktivovala lidem bez meditační zkušenosti kůru přední insuly. Míra aktivace odpovídá pravděpodobnosti odmítnutí nefér nabídky. Aktivace kůry přední insuly byla u lidí s meditační zkušeností v průběhu nefér nabídky nižší, zato se u nich ve větší míře aktivovala kůra zadní insuly. Ekonomicky racionální účastníci z kontrolní skupiny („lepší málo než vůbec nic“, například akceptovali víc než 85 % nespravedlivých nabídek) aktivovali dorsolaterální prefrontální kůru. Předpokládá se, že jde o projev růstu zátěže poznávacích funkcí. Ekonomicky racionální jedinci ze skupiny meditujících naproti tomu aktivovali somatosenzorickou a zadní horní spánkovou kůru. Při vyhodnocování nespravedlivé nabídky v průběhu hry Konec smlouvání, tedy lidé s meditační zkušeností, dokáží disociovat zápornou emoční odpověď od chování. Interocepce ovlivněná meditací tedy může ovlivňovat rozhodování.
Interocepce a empatie
Empatie umožňuje pochopit a sdílet pocity druhých lidí, je klíčová pro každodenní komunikaci a život v sociálním prostředí. Empatie je proces, v jehož průběhu prožíváme podobný senzorický, kognitivní a afektivní stav, jaký má člověk, kterého pozorujeme, přičemž si plně uvědomujeme, zda jsme zdrojem prožívání my sami, nebo pozorovaný jedinec. Podkladem empatického procesu je jak automatický afektivní prožitek, tak kontrolované kognitivní zpracovávání informací. Oba procesy aktivují rozličné neuronální systémy. Evolučně znamená empatie posilování sociálních vazeb. Umožňuje například pochopit, že je člen skupiny ve stresu. Empatie byla studována v řadě domén, příkladem jsou bolest, dotek nebo hnus. Neuronálním podkladem empatického procesu je činnost sítě tvořené oboustranně přední insulární kůrou, přední cingulární kůrou, thalamem a mediální prefrontální kůrou. Empatický proces je těžce narušen nebo zcela chybí u lidí s hraniční poruchou osobnosti, vývojovým autismem a u psychopatů (18).
Ernstová et al. (15) vyšetřovala vztah empatie a interocepce. Zdraví lidé sledovali standardizované emoční výrazy na fotografiích tváří Japonců a bělochů, žen i mužů. Fotografií bylo 56, zobrazovaly štěstí, smutek, hnus, strach, překvapení, hněv a opovržení. Pokusné osoby kromě toho sledovaly interoceptivní a exteroceptivní podněty, mlčky počítaly údery vlastního srdce a tóny. Empatická odpověď na pozorovanou tvář byla sledována funkční magnetickou resonancí tak, že empatická zátěž následovala buď po počítání srdečních úderů, nebo po počítání tónů, což se střídalo s kontrolní zátěží a „klidem“. Předcházející interocepce, v tomto případě pozornost věnovaná činnosti vlastního srdce, podstatně zvýšila aktivitu v průběhu empatického procesu v přední insulární kůře a některých korových oblastí ve střední čáře mozku. Interocepce je tedy podstatnou součástí empatického procesu.
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Thomayerova nemocnice
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Zdroje
1. Bartels A, Zeki S. The neural basis of romantic love. Neuroreport 2000; 11, p. 3829–3834.
2. Blood A. J., Zattore R. Intensely pleasurable response to music correlate with activity in brain regions implicated in reward and emotion. Proc Natl Acad Sci USA 2001, 98, p. 11818–11823.
3. Cauda F, D’Agata F, Sacco K., et al. Functional connectivity of insula in the resting brain. NeuroImage 2011;35, p. 8–23.
4. Craig AD. How do you feel? Interoception: the sense of the physiological condition of the body. Nat Rev Neurosci 2002; 3, p. 655–666.
5. Craig AD. Interoception: the sense of physiological condition of the body. Curr Opin Neurobiol 2003; 13, p. 500–505.
6. Craig AD. How do you feel-now? The anterior insula and human awareness. Nat Rev Neurosci 2009; 10, p. 59–70.
7. Critchley HD, Mathias CJ, Dolan RJ. Neuroanatomical basis for first and second-order representations of bodily states. Nat Neurosci 2001; 4, p. 207–211.
8. Critchley H. The human cortex responds to an interoceptive challenge. Proc Natl Acad Sci 2004; 101, p. 6333–6334.
9. Critchley HD, Wiens S, Rothestein, et al. Neural systems supporting interoceptive awareness. Nat Neurosci 2004; 7, p. 189–195.
10.Damasio A. The feeling of what happens: body and emotion in the making of consciousness. New York: Harcourt Brace, 1999.
11. Damasio A. Descartesův omyl. Emoce, rozum a lidský mozek. Praha: Kolumbus 2000.
12. Damasio A, Grabowski TJ, Bechara A, et al. Subcortical and cortical brain activity during the feeling of self-generated emotions. Nat Neurosci, 2000; 3, p. 1049–1056.
13. Damasio A. Fundamental feelings. Nature 2001; 413, p. 781.
14. Damasio A. Hledání Spinozy. Radost, strast a citový mozek. Praha: Dybbuk, 2003/2004.
15. Ernst J, Northoff G, Böker H, et al. Interoceptive awareness enhances neural activity during empathy. Hum Brain Mapp 2012; doi:10.1002/ hbm.22014.
16. Frith CD, Frith U. Interacting minds – a biological basis. Science 1999; 286, p. 1692–1695.
17. Kirk U, Downar J, Montague PR. Interoception drives increased rational decision-making in meditators playing the ultimatum game. Front Neurosci 2011; 5, p. 1–11.
18. Koukolík F. Základy kognitivní, afektivní a sociální neurovědy II. Empatie. Prakt. lék. 2011; 91, s. 63–67.
19. Koukolík F. Lidský mozek. Funkční systémy. Norma a poruchy. Třetí doplněné a rozšířené vydání. Praha: Galén, 2012.
20. Luders E, Toga AW, Lepore N, et al. The underlying anatomical correlates of long term mediation: larger hippocampal and frontal volumes of gray matter. NeuroImage 2009;45, p. 672–678.
21. Lutz A, Slagter HA, Dunne JD, et al. Attention regulation and monitoring in meditation. Trends Cogn Sci 2008; 12, p. 163–169.
22. Lutz A, Greischar LL, Perlman D, et al. BOLD signal in insula is differentially related to cardiac function during compassion meditation in experts vs. novices. NeuroImage 2009; 47, p. 1038-1046.
23. Motlová L., Koukolík F. Citový mozek. Neurobiologie, klinický obraz, terapie. Praha: Galén, 2006.
24. Mutschler T, Wieckhorst B, Kowalevski S, et al. Functional organization of anterior insular cortex. Neurosci Lett 2009, 26, p. 66–70.
25. Paus T. Primate anterior cingulate cortex: where motor control, drive and cognition interface. Nat Rev Neurosci 2001; 2, p. 417–424.
26. Pollatos O, Kirsch W, Chandry R. On the relationship between interoceptive awareness, emotional experience, and brain processes. Brain Res Cogn Brain Res 2005; 25, p. 948–962.
27. Tang YY, Ma Y, Fan Y, et al. Central and autonomic nervous system interactions is altered by short term meditation. Proc Natl Acad Sci 2009; 106, p. 8865–8870.
28. Websterův slovník: Random House Webster´s Unabridged Dictionary. 2nd edition. New York: Random House, 1998.
29. Weiss T, Straube T, Boettcher J et al. Brain activation upon selective stimulation of C-and A-δ fibers. NeuroImage 2008; 41, p. 1372–1381.
Štítky
Praktické lekárstvo pre deti a dorast Praktické lekárstvo pre dospelýchČlánok vyšiel v časopise
Praktický lékař

2012 Číslo 5
- Metamizol jako analgetikum první volby: kdy, pro koho, jak a proč?
- Fixní kombinace paracetamol/kodein nabízí synergické analgetické účinky
- Antidepresivní efekt kombinovaného analgetika tramadolu s paracetamolem
- Geriatrická křehkost a léčba bolesti
Najčítanejšie v tomto čísle
- Porucha chování v REM spánku – opomíjená diagnóza
- Eozinofilná pneumónia a systémová vaskulitída
- Atypický priebeh bronchopneumónie s extrapulmonálnymi komplikáciami spôsobenej infekciou Mycoplasma pneumoniae
- Významná aortální stenóza v běžné klinické praxi