
Anatomické a funkční vlastnosti extenzorů a hlubokých flexorů bérce, možnost využití při šlachovém transferu
Extensor and deep flexor muscles of the calf anatomical and functional properties, possibility of using in tendon transfer
INTRODUCTION:
This work describes the macroscopic fiber alignment of the anterior, lateral and deep posterior muscle groups of the calf, and skeletal muscle architecture. Latter is a basic factor, that determines muscle function. The most important parameters of muscle architecture are PCSA (Physiological crosssectional area), correlating with muscle strength, and Lf (muscle fiber length), which corresponds to the motion range of the muscles and tendons. Understanding these parameters and their relationship explains the physiologic basics of muscle strength and motion, and provides scientific evidence for muscle transfers.
MATERIALS AND METHODS:
In our study we analyzed 5 lower extremity preparates. On each, 8 muscles the anterior, lateral and deep posterior muscle groups of the calf were investigated. We measured and calculated the parameters of muscle architecture. We compared the characteristics of the individual muscles and muscle groups, and based on these results analyzed the possibilities of muscle transfer in the case of peroneal nerve injuries on various levels.
RESULTS:
The strongest of the investigated muscles is the posterior tibial muscle. The foot flexors have bigger muscle strength than the extensors. Supination forces acing on the foot are stronger than pronator forces. Musculus extensor hallucis longus has the biggest motion range. Extensors have bigger motion ranges than flexors.
CONCLUSION:
In case of isolated anterior tibial muscle lesions, transferring the long peroneal muscle to a neutral spot of the foot is the most favourable solution. In case of superficial peroneal nerve lesions, transfer of the long flexors of the toes to the tendons of the peroneal muscles is the best solution from muscle architectural point of view. The second option is transfer of the anterior tibial muscle. In case of profund peroneal nerve lesions, transfer of the posterior tibial muscle to the front side of the lower leg and medial side of the dorsum of the foot is the most advantageous. Regarding common peroneal nerve lesions, the only possibility is transferring the posterior tibial muscle on the front side of the lower leg to a neutral spot of the foot.
Key words:
muscle architecture, muscle fiber, tendon transfer, peroneal palsy.
Authors:
Petr Špiroch; Igor Čižmář; Jaromír Freiwald; Ján Palčák
Authors‘ workplace:
Department of Traumatology, University Hospital Olomouc, Czech Republic
; Traumatologické oddělení, Fakultní Nemocnice Olomouc, I. P. Pavlova 6, 775 20, Olomouc
Published in:
Úraz chir. 20., 2012, č.1
Overview
Úvod:
Práce popisuje makroskopické uspořádání svalových vláken svalů přední, laterální a zadní hluboké skupiny svalů bérce, architekturu kosterních svalů. Ta je základním faktorem určujícím funkci svalu. Nejdůležitějšími parametry svalové architektury jsou PCSA (Physiological crosssectional area), ukazatel svalové síly a Lf (délka svalových vláken), ukazatel rozsahu pohybu svalu a šlachy. Porozumění tomuto vztahu objasní fyziologický základ svalové síly a pohybu a poskytne vědecké odůvodnění operací šlachových transferů.
Materiál a metodika:
V experimentu bylo analyzováno 5 preparátů dolních končetin, na každém preparátu bylo analyzováno 8 svalů přední, laterální a zadní hluboké skupiny svalů bérce. Na svalech byly změřeny a vypočteny parametry svalové architektury. Byly porovnány hodnoty svalové architektury jednotlivých svalů a svalových skupin a na základě těchto hodnot byly rozebrány možnosti transferu při výpadku různých částí nervus peroneus.
Výsledky:
Nejsilnější ze zkoumaných svalů je musculus tibialis posterior. Flexory nohy mají větší svalovou sílu než extenzory. Supinační síly působící na nohu jsou větší než síly pronační. Největší rozsah pohybu má musculus extensor hallucis longus. Extenzory mají větší rozsah pohybu než flexory.
Závěr:
Při izolované lézi musculus tibialis anterior je nejvýhodnější transfer musculus peroneus longus do neutrálního bodu nohy. Pří lézi nervus peroneus superficialis je z pohledu svalové architektury nejvýhodnější transfer dlouhých flexorů palce a prstů na šlachy peroneálních svalů. Druhou možností je transfer předního tibiálního svalu. Při lézi nervus peroneus profundus je nejvýhodnější transfer zadního tibiálního svalu na přední stranu bérce a mediální stranu dorza nohy. Při lézi nervus peroneus communis je jediným možným transfer zadního tibiálního svalu na přední stranu bérce do neutrálního bodu nohy.
Klíčová slova:
architektura svalu, svalové vlákno, šlachový transfer, peroneální paréza.
Typ práce: Experiment.
Cíl práce:
Cílem práce je studium parametrů svalové architektury bércových svalů a rozbor výpadku funkce svalů při paréze nervus peroneus. Porovnáním parametrů svalové architektury svalů a svalových skupin určit optimální typ šlachového transferu k obnově ztracené funkce.
Úvod
Svalová soustava vytváří nervově řízený aktivní pohybový aparát. Základem je smrštění schopná příčně pruhovaná svalová tkáň, která je funkčně spjata s pohyblivě spojeným skeletem [7]. Základní stavební jednotkou svalu je svalové vlákno. Makroskopické uspořádání svalových vláken je nazýváno jako architektura kosterních svalů [12]. Protože architektura svalů je základním faktorem, určujícím funkci svalu, pochopení vztahu mezi strukturou a funkcí má velký praktický význam. Porozumění tomuto vztahu objasní nejen fyziologický základ svalové síly a pohybu, ale poskytne také vědecké odůvodnění operací šlachových transferů.
Anatomie
Svaly bérce dělíme do tří skupin, oddělených kostmi a vazivovými septy. Do přední skupiny patří extensory prstů a nohy, supinátory nohy musculus tibialis anterior (TA), musculus extensor digitorum longus (EDL), musculus extensor hallucis longus (EHL). Inervovány jsou n. peroneus profundus (NPP). Do laterální skupiny patří flexory a pronátory nohy musculus peroneus longus (PL) a musculus peroneus brevis (PB). Inervovány jsou n. peroneus superficialis (NPS). Zadní skupinu dělíme na povrchovou a hlubokou. Do povrchové vrsty patří musculus triceps surae, m. plantaris, do hluboké vrsty řadíme m.popliteus, m. tibialis posterior (TP), m. flexor digitorum longus (FDL) a m. flexor hallucis longus (FHL). Funkcí těchto svalů je flexe nohy a prstů a supinace nohy. Inervovány jsou n. tibialis [7].
Základní architektonické pojmy
Je publikováno množství článků popisujících sval na mikroskopické úrovni s mikrofotografiemi a schématy svalových sarkomer [5, 6, 20]. O uspořádání svalových vláken uvnitř a mezi svaly je publikováno mnohem méně. Architektura svalových vláken může být definována jako uspořádání svalových vláken uvnitř svalu vzhledem k ose sil [17]. Počet architektonických uspořádání svalů je stejný jako počet svalů samotných. Z praktického hlediska se svaly dělí do 3 až 4 obecných typů architektury svalových vláken.
Prvním typem jsou svaly složené z vláken, která jsou uspořádána rovnoběžně s osou sil; tato architektura je nazývána jako souběžná, nebo podélná. Příkladem tohoto typu je m. biceps brachii. Ostatní svaly jsou označovány jako svaly pennatní. Tento pojem vychází z latinského pinnatus pernatý, opeřený, okřídlený nebo vějířovitý. Ty se dělí na svaly, jejichž vlákna jsou orientována v jednom úhlu vzhledem k ose síly; tyto svaly jsou nazývány jako svaly unipennatní. Příkladem je m. vastus medialis a lateralis. Úhel vláken je měřen v klidovém stavu svalu. Z disekcí svalů je zřejmé, že většina svalů spadá do nejobecnější kategorie, multipennatních svalů, které jsou tvořeny vlákny svírajícími různé úhly k ose síly. Příkladem je např. m. gluteus maximus. Některá literatura [1] uvádí ještě typ bipennatní, u nějž jdou svalová vlákna ze dvou stran k ose pohybu. Nejpřesnějším příkladem tohoto typu je sval adduktoru klepete humra, u člověka se mu nejvíce podobá m. rectus femoris.
První práce zabývající se studiem a popisem architektoniky svalů byla publikována v roce 1965 Gansem a Bockem [12], kde byla precizně popsána metodika studia svalové architektury. Tato metodika byla upravena v roce 1987 Gansem a De Vriesem [13]. V těchto pracích byly definovány parametry, sloužící k analýze svalové architektury.
Prvním parametrem je délka svalu Lm a je definována jako vzdálenost od úponu nejproximálnějšího svalového vlákna k úponu nejdistálnějšího vlákna na šlachu. Délka svalového vlákna, nebo fasciklu Lf je měřena po mikrodisekci svalu od proximálního konce fasciklu k jeho úponu na šlachu. Dalším parametrem je hmotnost svalu M; jde o čistou hmotnost svalu bez šlachy.
Materiál a metoda
K výzkumu byly použity preparáty pěti dolních končetin. Při měření a analýze preparátů bylo postupováno dle metodiky popsané Gansem a De Vriesem [13]. Jednalo se o čerstvé nefixované preparáty dolních končetin od poloviny stehna distálně včetně prstů.
Odstranili jsme povrchové svaly a fascii svalů jak na bérci, tak na noze s ponecháním retinakul. Provedli jsme maximální flexi a pronaci nohy a prstů, označili jsme na šlachách TA, EHL a EDL místo v úrovni horního okraje retinaculum mm. extensorum superior, převedli jsme nohu do maximální supinace a extenze včetně prstů a udělali jsme na šlachách značku v tomto postavení nohy. Změřili jsme vzdálenost na šlachách, která určuje možnost maximálního pohybu daného svalu a šlachy. Hodnota je označena jako Rm, měřena s přesností 0,1cm. Obdobně jsme označili v extenzi a supinaci nohy šlachy m.peroneus longus a brevis při horním okraji horního peroneálního retinakula, převedli jsme nohu do flexe a pronace a změřili jsme rozsah pohybu peroneálních svalů a šlach. Poté jsme v extenzi a pronaci nohy označili šlachy TP, FDL a FHL v úrovni retinaculum m. flexorum, převedli nohu do flexe a supinace a změřili rozsah pohybu těchto svalů a šlach.
Následně jsme protnuli retinakula. Odetnuli jsme distální úpony šlach, svaly jsme ostře odpreparovali od periostu a od membrana interossea cruris. Jednotlivé svaly jsme rozprostřeli, na povrchu osušili, změřili jsme délku svalu Lm, s přesností na 0,1cm od úponu nejproximálnějších vláken k úponu nejdistálnějších vláken ke šlaše. Dalším měřeným parametrem je délka mimosvalové šlachy LT, kterou jsme měřili od úponu nejdistálnějšího svalového vlákna ke šlaše ke konci šlachy s přesností na 0,1cm. Svalovou část jsme rozměřili do tří třetin; uprostřed každé třetiny jsme vypreparovali svalový fascikl a změřili jsme délku svalového vlákna, fasciklu, Lf13, digitálním laserovým měřidlem s přesností na 0,02 cm. Z těchto tří naměřených hodnot byl vypočítán aritmetický průměr. Ve stejných místech jsme změřili standardním úhloměrem úhel, který svírají svalová vlákna k ose pohybu, distální šlaše pennation úhel s přesností na 0,5o, který je nazván Alfa13. Z naměřených hodnot byl vypočítán aritmetický průměr (obr. 1).
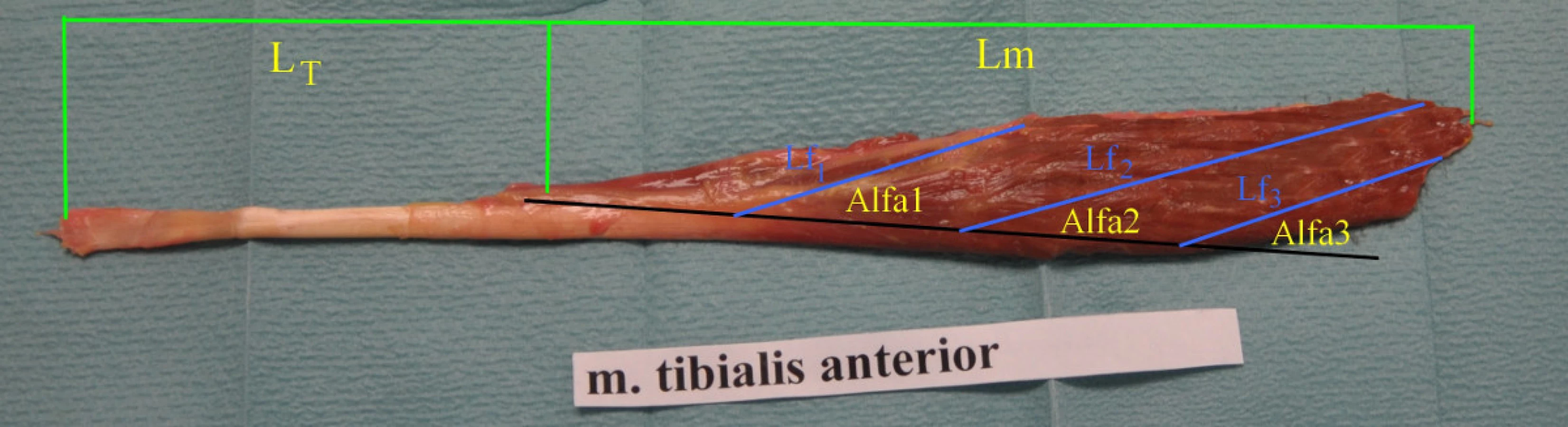
Z naměřených hodnot jsme vypočítali poměr mezi délkou svalových vláken k délce svalu (poměr Lf/Lm), dle popsané metodiky [13]. Tento poměr ukazuje možný rozsah pohybu svalu. Například, jestliže sval obsahuje vlákna, která probíhají celou délkou svalu (poměr Lf/Lm=1) je uzpůsoben k většímu rozsahu pohybu než sval, jehož vlákna mají jen poloviční délku k délce svalu (Lf/Lm=0,5). Tento poměr je dobře použitelný k porovnání rozsahu pohybu svalů, protože je nezávislý na absolutní délce svalových vláken a umožňuje porovnání rozsahu pohybů mezi svaly.
Druhý vypočítávaný parametr určuje maximální možnou sílu svalu [23]. Je nazýván PCSA (Physiological crosssectional area). Vypočítává se podle vzorce:
PCSA(cm2)= M(g)x cos Alfa/ p(g/cm3)xLf(cm) [2], kde M je hmotnost svalu bez šlachy, Alfa je povrchový pennation úhel, p je hustota svalu (u savčího svalu je tato hodnota 1,056g/cm3 [26]). Teoreticky PCSA představuje součet průřezů všech svalových vláken ve svalu. Tato rovnice vypočítává objem svalu (hmotnost/hustota) rozdělenou do svalových vláken a je udáván v plošných jednotkách (v tomto případě cm2). Protože jsou svalová vlákna orientována v určitém úhlu k ose pohybu, je do rovnice začleněn cosinus tohoto úhlu, čímž je zohledněno to, že ne všechna síla svalových vláken je převáděna na šlachu. Tato myšlenka vychází z modelu svalového vlákna, kdy je určitou silou taženo za šlachu v různých úhlech k ose pohybu. Výsledná síla tahu za šlachu je vždy rovna síle x cosinus úhlu tahu. To znamená, že čím větší je úhel, tím menší je převáděná síla a pokud dosáhne úhel 90°, je výsledná síla 0.
Výsledky
Celkem jsme analyzovali na 5 preparátech 40 svalů, u nichž byly měřeny a vypočítány parametry svalové architektury popsané v metodice (tab. 1). Dvěma nejdůležitějšími parametry svalové architektury jsou PCSA, jakožto ukazatel svalové síly a délka svalových vláken, respektive poměr délky svalových vláken k délce svalu, jakožto ukazatel rozsahu pohybu šlachy a svalu. Když zaneseme zjištěné hodnoty pro jednotlivé svaly do bodového grafu, kdy délka svalových vláken je na ose X a PCSA na ose Y, získáme přehled o podobnosti nebo rozdílnosti síly a pohybu jednotlivých svalů, a tím i vhodnosti ke šlachovým transferům.
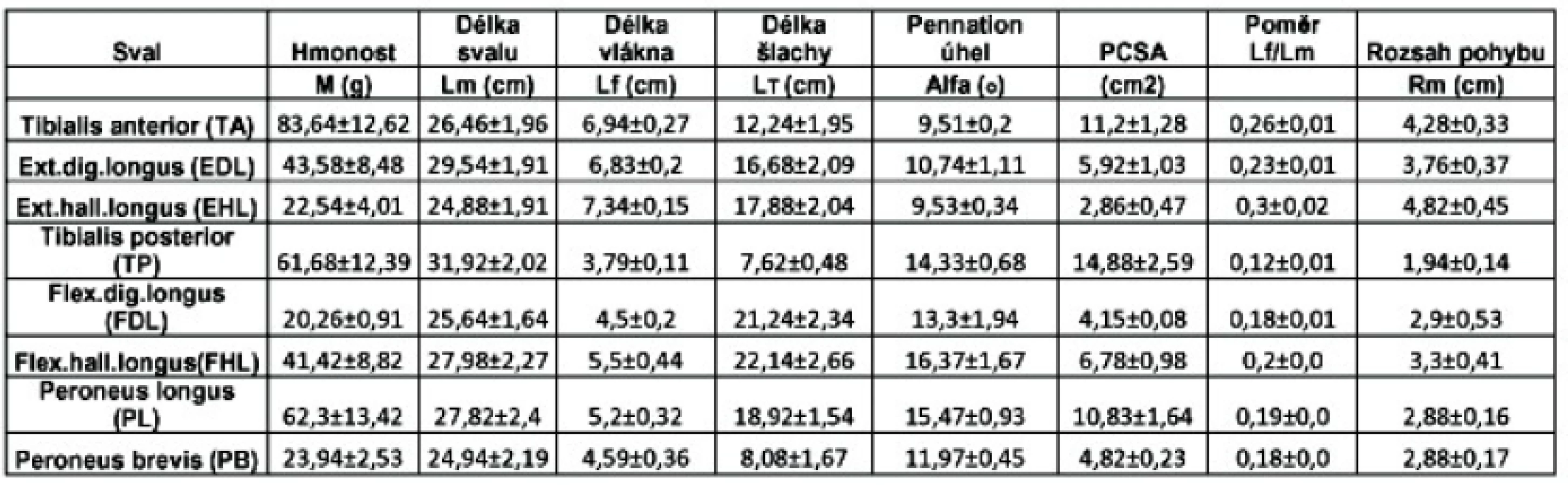

Z výsledků vyplývá, že nejsilnějším zkoumaným svalem (na základě PCSA) je TP. Druhým nejsilnějším svalem je TA. Ze součtu PCSA jednotlivých svalových skupin vyplývá, že flexory nohy a prstů jsou silnější než extenzory. Supinátory nohy (TP+TA) jsou silnější, mají 62 % svalové síly, než její pronátory (PL+PB), 38 % svalové síly. Z extenzorů připadá 56 % svalové síly extenze na TA, 30 % na EDL a 14 % na EHL, z čehož plyne, že při výpadku funkce TA je extenze nohy významně oslabena a EHL a EDL ji mohou nahradit je částečně a to za cenu hyperextenze prstů. Celkovou sílu flexorů nelze takto analyzovat, protože na flexi nohy se podílí další svaly m. triceps surae a m. plantaris, jejichž analýza nebyla součástí experimentu a jde o svaly, které se podílejí i na flexi kolene, nicméně 58 % svalové síly hlubokých flexorů připadá na TP, 26 % na FHL a 16 % na FDL. Při supinaci nohy 57 % svalové síly nohy připadá na TP a 43 % na TA. Pronaci nohy provádí z 69 % PL a z 31 % PB.
Největší změřený rozsah pohybu má EHL. Tomu odpovídá i to, že EHL má nejdelší svalová vlákna, jakožto ukazatel rozsahu pohybu, respektive největší hodnotu poměru délky svalových vláken k délce svalu. Pokud vezmeme jako základní délku pohybu měřený Rm TA a přepočteme ji podle poměru Lf/Lm na ostatní svaly dle vzorce Rm vypočtený= (Rm TA/0,26) x (Lf/Lm svalu) získáme vypočtený teoretický rozsah pohybu pro každý sval. Tento model výpočtu nebyl dle dostupné literatury aplikován, jde o vlastní invenci, umožňující srovnání měřených a předpokládaných hodnot při dané metodice. Při srovnání Rm měřeného a vypočteného není rozdíl statisticky významný, z čehož plyne, že rozsah pohybu je závislý na délce svalového vlákna, respektive na poměru délky svalového vlákna k délce svalu. Při porovnání rozsahu pohybu svalových skupin mají extenzory větší rozsah pohybu než flexory; nejpohyblivější šlachy jsou flexor a extenzor palce. Obě peroneální šlachy mají prakticky stejný rozsah pohybu, což odpovídá jejich stejnému průběhu a stejné funkci.
Klinická aplikace výsledků experimentu
Při výpadku funkce NPP jsou postiženy všechny svaly přední svalové skupiny, při výpadku funkce NPS svaly laterální svalové skupiny. Při postižení n. peroneus communis (NPC) je výpadek funkce obou těchto svalových skupin. V klinické praxi je také významné postižení samotného TA, a to z inervačních poruch, nebo při lézích samotného svalu. Pro potřeby biomechanických studií hlezna a nohy a šlachových tranferů byl definován tzv. neutrální bod nohy, kterým je na dorzu nohy os cuneiforme laterale a baze III. metatarzu, kdy při tahu za toto místo do extenze nedochází ani k pronaci, ani supinaci nohy. Při přenosu na mediální stranu nohy se transferovaná šlachy přenáší na os cuneiforme mediale, nebo bazi I. metatarzu. Při přenosu na zevní stranu nohy se transferovaná šlacha přenáší na bazi 5. metatarzu. Ztráta síly flexe při transferu jednotlivých svalů je nevýznamná, proto není analyzována. Tato analýza je matematická, na základě parametrů svalové architektury a nebere v úvahu další faktory, které se mohou podílet na výsledné svalové síle a rozsahu pohybu.
Rozbor výpadku funkce musculus tibialis anterior
Při výpadku funkce samotného TA přichází noha o 56 % svalové síly extenze a o 43 % supinace. Tím dojde k vyrovnání až k lehké převaze pronačních sil na nohu (51 % pronace, 49 % supinace). Svaly s dostatečnou silou, na základě PCSA, které mohou funkci TA (PCSA 11,2 cm2) nahradit, jsou PL (PCSA 10,83 cm2) a TP (PCSA 14,88 cm2). Při transferu PL do neutrálního bodu nohy dochází k převaze supinačních sil, a to 76 % supinace, která je zajišťována TP a 24 % pronace, kterou zajišťuje PB (graf 2). Při transferu PL na přední stranu bérce a mediální stranu nohy je převaha supinačních sil již významná (84 %). Při transferu TP na přední stranu bérce a mediální stranu nohy zůstává poměr sil pronace a supinace vyrovnaný (51 % pronace, 49 % supinace). Při transferu TP do neutrálního bodu nohy by byla supinační síla nulová a pronační 100 %. Celková délka svalu a šlachy TP je srovnatelná s délkou TA, PL má díky průběhu plantou delší šlachovou část. TA má velikost pohybové exkurze 4,28 cm, TP 1,94 cm a PL 2,88 cm.

V tomto případě jsou supinační síly převažující (70 %), pronace (30 %) (graf 3).

Rozbor výpadku funkce n. peroneus spfc:
Při poškození NPS dochází k výpadku funkce PL a PB, tedy obou pronátorů nohy. K náhradě pronace nohy mohou sloužit prakticky všechny ostatní funkční svaly, které byly popsány. Při transferu flexorů zůstanou peroneální šlachy ve svém původním uložení za zevním kotníkem. Transfer TP na peroneální šlachy způsobí převahu pronačních sil (57 %) proti supinačním silám (43 %). Aby měly flexory palce a prstů dostatečnou sílu, je potřeba je přenést společně.
Při transferu šlach přední skupiny svalů bérce musí šlachy probíhat na dorzu hlezna a nohy, před fibulou, aby neztratily svou extenční funkci. Při transferu TA jsou supinační síly 57 % a pronační 43 %. Extenzory prstů a palce musí být také přenášeny společně, aby měly dostatečnou sílu. Pak je pronace 25 % a supinace 75 %. Flexory a extenzory palce a prstů jsou výhodnější pro delší šlachové části proti TA a TP. Flexory prstů a palce mají nejpodobnější rozsah pohybu (2,9 cm FDL a 3,3 cm FHL) jako peroneální šlachy (2,88 cm). TP má malý rozsah pohybu (1,94 cm), extenzory výrazně větší (TA 4,28 cm, EDL 3,76 cm, EHL 4,82 cm).
Rozbor výpadku funkce n. peroneus profundus
Při poškození NPP dochází k výpadku funkce TA, EHL a EDL a také krátkých extenzorů prstů. Není tedy možná aktivní extenze prstů a nohy a je mírná převaha pronačních sil (51 %). K náhradě extenční funkce je potřeba se co nejvíce přiblížit PCSA přední svalové skupiny, které je 19,98 cm2.
Ten je potřeba přenést na přední stranu bérce a mediální stranu dorza nohy k udržení supinační funkce svalu (49 %) (graf 4). Při jeho přenosu do neutrálního bodu nohy by byly supinační síly nulové. Nevýhodou TP je krátký rozsah pohybu (1,94 cm oproti TA 4,28 cm), který vede k omezení flexe nohy a hraniční délka svalu a šlachy (TA 38,7 cm, TP 39,54 cm) k transferu. K posílení extenze nohy a obnově aktivní extenze prstů je možné přenést spolu s TP také šlachu FDL (PCSA TP+FDP 19,03 cm2), nebo FHL (PCSA TP+FHL 21,66 cm2) na šlachy EHL a EDL. Další možností je přenos PL (PCSA 10,83 cm2) na přední stranu bérce do neutrálního bodu nohy (supinační síly 76 %, pronační 24 %), nebo na mediální stranu nohy (supinace 84 %, pronace 16 %), spolu s FDL (PCSA PL+FDL 14,98 cm2), nebo FHL (PCSA PL+FHL 17,61 cm2), nebo s oběma flexory prstů (PCSA PL+FDL+FHL 21,76 cm2) na extenzory palce a prstů. PL má podobnější rozsah pohybu (2,88 cm) jako TA a delší celkovou délku (46,74 cm).
Rozbor výpadku funkce n. peroneus communis
Při poškození NPC dochází k výpadku funkce TA, EHL, EDL, PL a PB. Funkční zůstávají jen TP, FHL a FDL. Noha je v supinačním postavení tahem TP. Svalem s dostatečnou silou k obnově aktivní extenze je TP, který je potřeba přenést na přední stranu bérce do neutrálního bodu nohy k vyřazení jeho supinační aktivity, která by při přenosu na mediální stranu nohy zůstala. V tomto případě na nohu nepůsobí ani pronační ani supinační síly. Spolu s TP je možné přenést FDL na extenzory palce a prstů (PCSA TP+FDP 19,03 cm2) nebo FHL na extenzory prstů a palce (PCSA TP+FHL 21,66 cm2), čímž je obnovena extenze prstů a palce a zůstává zachována flexe prstů a palce. Malým rozsahem pohybu TP (1,94 cm) oproti TA (4,28 cm) dochází k omezení flexe hlezna.

Diskuze
Hodnoty svalové architektury jsou nejlepším ukazatelem funkce svalů [18]. Muskuloskeletální modely vycházejí z těchto parametrů. Chirurgické postupy při šlachových transferech, jejichž počátky jsou ve 20. a 30. letech 20. století, původně vycházely z anatomických modelů a empirie, snahou nahradit funkci poškozeného svalu, bez bližších znalostí svalové architektury. Později, od 70. let 20. století, ve kterých jsou publikovány první práce zabývající se svalovou architekturou, jsou tyto postupy buď obhájeny [16, 27], když přenášený sval je nejvýhodnější i z hlediska architektonických parametrů svalů, nebo jsou postupy měněny zjištěním, že je možné použít rozsahem pohybu nebo svalovou silou výhodnější sval [3, 19, 23]. Týká se to především horní končetiny, předloktí, kde je více svalů a tím více možností šlachových transferů, a to hlavně zásluhou Jana Fridéna, který v posledních dvou dekádách publikoval nejvíce prací týkajících se svalových studií a šlachových transferů [9, 10, 11].
V tomto experimentu bylo cílem věnovat se svalům bérce, vhodným ke šlachovým transferům, při výpadku funkce svalů inervovaných n. peroneus. Dále určení parametrů svalové architektury, svalové síly, pohybových exkurzí, popisu rozdílů mezi jednotlivými svalovými skupinami, určením podílu svalové síly jednotlivých svalů při extenzi, pronaci a supinaci nohy a z těchto parametrů určit nejvýhodnější svaly k transferu a místa, na která mají být přeneseny.
Jsou ovšem také určité faktory, které limitují přesnost a možnost interpretace zjištěných dat. Jedním z nich je pravděpodobně vyšší věk zemřelých jedinců, ze kterých pochází svalové preparáty, což vede vzhledem k atrofii svalů a nižší svalové síle k nižšímu zjištěnému PCSA jakožto determinantu svalové síly, než by bylo zjištěno u mladších jedinců [25]. To je problém absolutních hodnot PCSA. Při srovnávání svalových skupin jsou poměrné výsledky správné pro předpoklad symetrické atrofie všech svalů a svalových skupin. Dalším faktorem, který může ovlivnit délku svalu a délku svalového vlákna, je poloha kloubu v okamžiku jeho fixace [14]. Předpokládá se fixace v neutrálním postavení v kloubu. I když je to opět problém jen interpretace absolutních hodnot, při použití poměrných hodnot, délky svalového vlákna k délce svalu, jsou výsledky srovnatelné. Tuto teorii podporuje i náš experiment, kdy srovnáním hodnot svalové architektury v podobných pracích, jsou absolutní hodnoty rozdílné, ale poměrné a vypočtené hodnoty podobné.
Přesnost této metody analýzy svalové síly byla ověřena na základě porovnání vypočítaného maxima svalové síly (založeného na PCSA) a měřeného maxima svalové síly (při použití tradičních fyziologických technik testování svalové síly) na svalech morčete [23]. Bylo zjištěno, že odhady a předpovědi svalové síly souhlasily s experimentálně naměřenými hodnotami, s výjimkou m. soleus. Pro všechny tzv. „rychlé“ svaly byla změřena svalová síla na jednotku průřezu 22,5 N/ cm2, která slouží jako jmenovitá jednotka napětí savčích svalů.
Metoda výpočtu svalové síly předpokládá, že je pennation úhel v průběhu kontrakce svalu konstantní. Experimentálním měřením pennation úhlu na svalových vláknech m. gastrocnemius u krysy, který je unipennátní, bylo zjištěno, že tomu tak není [30]. Zuurbier a Huijing [29] umístili malou drátěnou značku přes povrch m. gastrocnemius a natáčeli pohyb značky během svalové kontrakce. Za použití těchto dat změřili pennation úhel vláken a ukázali, že se úhel vláken značně liší se změnou délky svalu během jeho fyziologického pohybu. Když byl klidový úhel vlákna asi 30°, během izotonické kontrakce se úhel zvýšil na téměř 60°. A svalová aponeuroza se k ose svalu otáčí o 10–15°. Skutečnost, že se svalová vlákna mohou během kontrakce volně otáčet má své důsledky. Prvním je, že vypočítané PCSA předpovídá sílu jen pro jednu polohu svalu a má omezenou výpovědní hodnotu pro předpověď síly v průběhu celé kontrakce. Druhým je, že rotace svalových vláken má významný dopad na odhad rychlosti svalových vláken na základě měření rychlosti celých svalů. Když vezmeme m. gastrocnemius laboratořního potkana, a změříme rychlost kontrakce svalového vlákna při udržení konstantního pennation úhlu, bude se jednat také o rychlost kontrakce celého svalu. Pokud ovšem budeme počítat s rotací vláken, tak je rychlost kontrakce jednotlivých vláken ve skutečnosti značně nižší než rychlost kontrakce celých svalů. Takto rotace svalových vláken během kontrakce svalu zajistí svalovým vláknům možnost větší produkce síly, než kdyby zůstávala v konstantním pennation úhlu. Reeves [24] ve své ultrazvukové studii in vivo popisuje svalovou architekturu m. tibialis anterior během pohybu. Ve své práci popisuje stav svalu při izometrické kontrakci, kdy zjistil, že během izometrické kontrakce bylo svalové vlákno kratší a pennation úhel větší v porovnání s klidovými hodnotami. Dále měřil pennation úhel při pohybu v hleznu při různé rychlosti pohybu. Během izotonické koncentrické kontrakce od 0 do 4,36rad/s stoupá délka vláken, měřená v konstantním úhlu hlezna z 49,5 mm na 69,7 mm a pennation úhel klesá z 14,8° na 9,8°. Během excentrické kontrakce se fascikly stahují jakoby izometricky, nezávisle na úhlové rychlosti. Toto chování svalu během kontrakce je v důsledku působení napětí na elastické komponenty svalu, které klesá se stoupající rychlostí kontrakce. Pseudoizometrické chování fasciklů během excentrického pohybu svalu je tedy způsobeno řadami elastických komponent ve svalu, které vyrovnávají excentrické pohyby.
I když je často uváděno, že délka svalového vlákna je přímo úměrná pohybovému rozsahu vlákna (nebo jeho rychlosti), nebyla publikována žádná ucelená studie savčích svalů analogická studiím PCSA (vypočteného a zjištěného měřením). Nicméně v literatuře je k dispozici hodně experimentálních důkazů, že tento vztah je platný. Například Brand [4] a Woittiez [28] dokazují v mechanické studii na izolovaných žabích svalových vláknech, změřením délky vlákna, počtu sarkomer v řadě ve vláknu, a změřením rychlosti a rozsahu pohybu, že maximální rychlost kontrakce je přímo úměrná délce vláken a podobný je i vztah při izometrické kontrakci mezi délkou a napětím vlákna. Toto je vlastně důvod, proč bývá rychlost kontrakce označována jednotkou délky vlákna/s, nebo délky sarkomery/s. Jiným důkazem jsou anatomickomechanické studie m. semitendinosus u kočky, kozy a morčete [2, 8, 14, 15, 23]. Musculus semitendinosus poskytuje unikátní model svalu, který je složen ze samostatné proximální a distální hlavy, které jsou odděleny šlachou a každá z nich má samostatnou inervaci. Bylo zjištěno, že při stimulaci obou hlav současně je maximální rychlost stahu stejná jako součet maximálních rychlostí při stimulaci hlav samostatně. V souhlasu s těmito pracemi je i zjištění v této studii, že při srovnání rozsahu pohybu změřeného (Rm měřený) a vypočteného na základě změření svalových vláken (fasciklů), vypočtení poměru k délce svalu a výpočtu teoretického rozsahu pohybu (Rm vypočtený), není rozdíl statisticky významný. Vztah mezi délkou svalového vlákna a délkou svalu může být komplikován zjištěním, že některé svaly s makroskopicky dlouhými svalovými vlákny (fascikly) jsou složeny z mnohem kratších svalových vláken, která jsou uložena v řadě a navazují na sebe [21, 22]. Tato skutečnost, spolu s elektromyografickými údaji, kdy za sebou seřazená svalová vlákna jsou inervována větvemi axonů, také uspořádanými v řadě, dává předpoklad, že všechna tato svalová vlákna budou aktivována současně, a tím budou funkčně působit jako jedno celé, dlouhé vlákno. Pokud by k současné aktivaci celé řady svalových vláken nedocházelo, mohla by nastat mechanicky značně nevýhodná situace, kdy při stahu periferní části svalu by proximální byla již relaxována.
Závěr
Z analýzy architektury svalů, rozsahu pohybu vyplývá, že při izolované lézi TA je nejvýhodnější transfer PL do neutrálního bodu nohy, čímž se zajistí dostatečná svalová síla a vyvážené pronosupinační síly působící na nohu.
Pří lézi NPS je z pohledu svalové architektury nejvýhodnější transfer FHL a FDL na šlachy peroneálních svalů, kdy je příznivý poměr pronačních a supinačních sil, podobný pohybový rozsah, dostatečná délka šlachových částí. Šlachy mohou probíhat za zevním kotníkem v původním uložení a tím mají nejfyziologičtější pohyb. Z hlediska poměru pronačních a supinačních sil, je možný transfer TA, který má kratší šlachu a větší rozsah pohybu.
Při lézi NPP je nejvýhodnější transfer TP na přední stranu bérce a mediální stranu dorza nohy spolu s FDL na extenzory prstů a palce. Tím je zajištěna dostatečná síla extenze nohy a prstů, vyrovnaný poměr pronačních a supinačních sil. Krátkým rozsahem pohybu TP dojde k omezení flexe nohy. Druhou možností je přenos PL do neutrálního bodu nohy spolu s přenosem FDL na přední stranu bérce a extenzory prstů a palce, kdy je sice nižší svalová síla, ale PL umožní větší rozsah pohybu hlezna.
Při lézi NPC je jediným možným transfer TP na přední stranu bérce do neutrálního bodu nohy. Spolu s TP je výhodné přenesení také FDL nebo FHL na extenzory palce a prstů k obnově jejich extenze a posílení extenze nohy.
MUDr. Petr Špiroch
email: Petr.Spiroch@fnol.cz
Sources
1. Azizi, E., Brainerd, EL., Roberts, TJ. Variable gearing in pennate muscles. Proc Natl Acad Sci USA. 2008, 105, 1745–1750.
2. Bodine, SC., Roy, RR., Meadows, DA. et al. Architectural, histochemical, and contractile characteristics of a unique biarticular muscle: the cat semitendinosus. J Neurophysiol. 1982, 48, 192–201.
3. Brand, PW. Tendon transfers for median and ulnar nerve paralysis. Orthop Clin North Am. 1970, 1, 447–454.
4. Brand, PW., Beach, RB., Thompson, DE. Relative tension and potential excursion of muscles in the forearm and hand. J Hand Surg Am. 1981, 3A, 209–219.
5. Burkholder, TJ., Lieber, RL. Sarcomere length operating range of vertebrate muscles during movement. J Exp Biol. 2001, 204, 1529–1536.
6. Burkholder, TJ., Lieber, RL. Sarcomere num-ber adaptation after retinaculum release in adult mice. J Exp Biol. 1998, 201, 309–316.
7. Čihák, R. Anatomie 1. Praha: Grada. 2001. p 313, 418–423. ISBN 80-7169-970-5
8. English, AWM., Weeks, OI. An anatomical and functional analysis of cat biceps femoris and semi-tendinosus muscles. Journal of Morphology. 1987, 191, 161–175.
9. Fridén, J., Lovering, RM., Lieber, RL. Fiber length variability within the flexor carpi ulnaris and flexor carpi radialis muscles: implications for surgical tendon transfer. J Hand Surg Am. 2004, 29, 909–914.
10. Fridén, J., Reinholdt, C. Current concepts in reconstruction of hand function in tetraplegia. Scand J Surg. 2008, 97, 341–346.
11. Fridén, J., Shillito, MC., Chehab, EF. et al. Mechanical feasibility of immediate mobilization of the brachioradialis muscle after tendon transfer. J Hand Surg Am. 2010, 35, 1473-1478.
12. Gans, C., Bock, WJ. The functional significance of muscle architecture: a theoretical analysis. Adv Anat Embryol Cell Biol. 1965, 38, 115–142.
13. Gans, C., De, Vries, F. Functional bases of fiber length and angulation in muscle. J Morphol. 1987, 192, 63–85.
14. Gans, C., Loeb, GE., de Vree, F. Architecture and consequent physiological properties of the semi-tendinosus muscle in domestic goats. Journal of Mor-phology. 1989, 199, 287–297.
15. Hutchison, DL., Roy, RR., Bodine-Fowler, S. et al. Electromyographic (EMG) amplitude patterns in the proximal and distal compartments of the cat semitendinosus during various motor tasks. Brain Res. 1989, 479, 56–64.
16. Lieber, RL. Skeletal muscle architecture: implications for muscle function and surgical tendon transfer. J Hand Ther. 1993, 6, 105–113.
17. Lieber, RL. Skeletal muscle structure and function: implications for physical therapy and sports medicine. Baltimore: Williams & Wilkins. 1992. pp. 303.
18. Lieber, RL., Brown, CC. Quantitative method for comparison of skeletal muscle architectural proper-ties. J Biomech. 1992, 25, 557–560.
19. Lieber, RL., Fazeli, BM., Botte, MJ. Architecture of selected wrist flexor and extensor muscles. J Hand Surg. 1990, 15A, 244–250.
20. Lieber, RL., Fridén, J. Functional and clinical significance of skeletal muscle architecture. Muscle Nerve. 2000, 23, 1647–1666.
21. Loeb, GE., Pratt, CA., Chanaud, CM. et al. Distribution and innervation of short, interdigitated muscle fibers in parallel-fibered muscles of the cat hind limb. J Morphol. 1987, 191, 1–15.
22. Ounjian, M., Roy, RR., Eldred, E. et al. Physiological and developmental implications of motor unit anatomy. J Neurobiol. 1991, 22, 547–559.
23. Powell, PL., Roy, RR., Kanim, P. et al. Predictability of skeletal muscle tension from architec-tural determinations in guinea pig hind limbs. J Appl Physiol. 1984, 57, 1715-1721.
24. Reeves, ND., Narici, MV. Behavior of human muscle fascicles during shortening and lengthening contractions in vivo. J Appl Physiol. 2003, 95, 1090–1096.
25. Ward, SR., Eng, CM., Smallwood, LH. et al. Are current measurements of lower extremity muscle architecture accurate? Clin Orthop Relat Res. 2009, 467, 1074–1082.
26. Ward, SR., Lieber, RL. Density and hydration of fresh and fixed skeletal muscle. J Biomech. 2005, 38, 2317–2320.
27. Wickiewicz, TL., Roy, RR., Powell, PL. et al. Muscle architecture of the human lower limb. Clin Orthop. 1983, 179, 275–283.
28. Woittiez, RD., Baan, GC., Huijing, PA. et al. Functional characteristics of the calf muscles of the rat. J Morphol. 1985, 184, 375–387.
29. Zuurbier, CJ., Huijing, PA. Changes in geometry of actively shortening unipennate rat gastrocnemius muscle. J Morphol. 1993, 218, 167–180.
30. Zuurbier, CJ., Huijing, PA. Influence of muscle geometry on shortening speed of fibre, aponeurosis and muscle. J Biomech. 1992, 25, 1017–1026.
Labels
Surgery Traumatology Trauma surgeryArticle was published in
Trauma Surgery

2012 Issue 1
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
- Metamizole at a Glance and in Practice – Effective Non-Opioid Analgesic for All Ages
- Metamizole vs. Tramadol in Postoperative Analgesia
- Spasmolytic Effect of Metamizole
- Safety and Tolerance of Metamizole in Postoperative Analgesia in Children
Most read in this issue
- ORIF (otvorená repozícia a interná fixácia) u dislokovaných intraartikulárnych zlomenín kalkanea, 2. - praktická časť
- Chyby a komplikace při hřebování zlomenin proximálního femuru hřebem TARGON PF
- Anatomické a funkční vlastnosti extenzorů a hlubokých flexorů bérce, možnost využití při šlachovém transferu
- Damage control laparotomie u úrazového hemoperitonea