
Základy kognitivní, afektivní a sociální neurovědy
III. Agrese a násilné chování
Basics of social cognitive and affective neuroscience; III. Aggression and violence
The nature of human aggression can be adressed by considering of Tinbergen’s four ethological questions:
- functional explanation,
- phylogenetic origin,
- developmental origin and modification through experience, and
- mmediate causes.
Is homicidium an adaptation or is it not? What is a function of homicidal fantasies?
Phylogenetic origins of aggression is tied to its evolutionary function. Aggression can be observed throughout the animal kingdom. The neural circuits that underlie aggressive behaviour lie in the forebrain, midbrain and hindbrain.
Key words:
aggression, evolution, ethology, phylogenesis.
Autoři:
F. Koukolík
Působiště autorů:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
Vyšlo v časopise:
Prakt. Lék. 2011; 91(3): 123-126
Kategorie:
Editorial
Souhrn
Povahu lidské agrese vysvětlují odpovědi na čtyři Tinbergenovy etologické otázky:
- funkční význam,
- fylogeneze,
- ontogeneze spolu s modifikacemi podmíněnými učením, a
- bezprostřední příčiny.
Je homicidium adaptace, nebo není? Jaký je význam homicidních fantazií?
Fylogenetický původ agrese je ve vazbě na její evoluční funkci. Agresi lze pozorovat v celé živočišné říši. Neuronální obvody, jejichž činnost je podkladem agrese, jsou v telencefalu, mesencefala i rhombencefalu.
Klíčová slova:
agrese, evoluce, etologie, fylogeneze.
Úvod
U zvířat se popisuje dvojí typ agrese: na jedné straně agrese označovaná jako defenzivní, reaktivní, hostilní, afektivní akt, na straně druhé jako akt ofenzivní, dravčí, promyšlený, instrumentální (3; přehled 28). Dichotomické členění tohoto druhu však zjednodušuje.
Klasifikace agrese zvířat uvádí osm typů (23, 30):
- cílená na predátory
- defenzivní, navozená strachem
- predátorní
- agrese mezi samci týkající se dominance
- mateřská při ochraně mláďat
- agrese vázaná na sexuální chování
- teritoriální
- navozená šokem
Odhaduje se, že v současnosti žije na Zemi asi 10 milionů živočišných druhů, 4 000 druhů z nich jsou savci.
Mezi savci pouze dva druhy tvoří koalice, které pořádají koordinované nájezdy do území obývaných jinými skupinami příslušníků vlastního druhu s cílem zabíjet:
- šimpanzi, a
- lidé.
U lidí se za agresi se považuje jakékoli chování cílené na druhého člověka s bezprostředním záměrem ublížit. Pachatel přitom musí být přesvědčen, že „cíli“ ublíží a cíl je motivován k tomu, aby se agresi vyhnul. Násilné chování (violence) je agrese, jejímž cílem je krajní poškození oběti, například její smrt (1; přehled 19).
Všechny případy násilného chování jsou agrese, mnohé případy agrese však nejsou násilné chování. Jestliže jedno tříleté dítě shodí druhé stejně staré dítě z tříkolky proto, aby ji mělo pro sebe, jde o agresi, o násilné chování se nejedná.
Světová zdravotnická organizace (31) definuje násilné chování (violence) slovy „záměrné užití fyzické síly nebo moci, buď jako hrozby, nebo aktuálně, a to proti sobě samému, druhé osobě, proti skupině či komunitě, které buď s vysokou pravděpodobností vyústí, nebo skutečně vyústí do úrazu, smrti, psychologického poškození, naruší vývoj nebo způsobí deprivaci.“
Definice zahrnuje násilné chování mezi osobami včetně ozbrojených konfliktů stejně jako sebevražedné chování, týká se i zastrašování a ohrožování.
Jak se agrese a násilné chování vysvětlují?
Teorie mluvící o „zabijáckém instinktu“ odvozovaná u učení Sigmunda Freuda a Konráda Lorenze mnoho neobjasňuje. Pojmenování není vysvětlením. Plodnější je chápat agresi jako řešení většího počtu různých druhů adaptivních problémů (7, 13).
Prvním z nich je získání zdrojů druhých lidí, například
- zemědělské půdy,
- potravin,
- nástrojů,
- zlata a šperků,
- zbraní,
- plodných žen,
- přístupu k pitné vodě.
Druhým je obrana proti útoku.
Třetím problémem jsou rivalové stejného pohlaví soutěžící o stejný zdroj, jímž jsou cenění příslušníci druhého pohlaví.
Problémem čtvrtým je vzestup na mocenském žebříčku skupiny.
Pátým, šestým a sedmým důvodem agrese jsou odrazování sexuálního partnera od nevěry, pokus získat ztraceného pohlavního partnera, jakož i zisk sexuálního přístupu k partnerovi nebo partnerce, které jinak získat nelze.
Definice agrese
Etologicky lze agresi zvířat a lidí popsat odpověďmi na čtyři Tinbergenovy otázky (27):
- jaký je její adaptivní význam, jak přispívá k přežití jedince a jeho naději na reprodukci;
- jaká je její fylogeneze
- jaká je její ontogeneze a modifikace zkušeností neboli učením
- jaké jsou její bezprostřední příčiny
Adaptivní význam
Agrese mezi zvířaty se vysvětluje soutěží o zdroje a ochranou mláďat. Míra agrese je proměnlivá a je úměrná míře dostupnosti zdrojů potravy a míře dostupnosti sexuálních partnerů, s nimiž je možné mít potomky. Teoretickým principem je analýza poměru nákladů a prospěchu (C/B, cost/ benefit). Jestliže jsou náklady na získání potravy větší, než je prospěch, zvířata nemusí hájit teritorium, jestliže je dostupnost samiček malá, chovají se samečkové útočněji.
Evoluce agrese byla studována pomocí teorie her. Interakci různých strategií lze vypočítat „výplatní“ maticí (pay-off matrix). Maynard-Smith (22) tímto způsobem formuloval takzvanou evolučně stabilní strategii, což je strategie, kterou v dané populaci s ohledem na poměr nákladů a prospěchu nelze nahradit strategií alternativní.
Modely, které při studiu agrese zvířat užily teorii her, vysvětlují, proč je výsledkem přírodního výběru útok, který soka spíše zastrašuje a odhání, než by ho zabil. Řada druhů zvířat je s to vyhodnotit bojový potenciál soka dřív, než se do boje pustí, například na základě sokovy velikosti, takže ustoupí a střet nenastane.
Lze lidskou schopnost vraždit chápat jako adaptaci?
Duntleyova a Bussova teorie vraždy jako adaptace, neboli homicidní adaptivní teorie (7,13, 14), předpokládá, že se u lidí, u mužů víc než u žen, vyvinul mechanismus umožňující vraždu příslušníka vlastního druhu v okolnostech, které lze předpovědět, například v průběhu války, sexuální rivality a partnerské nevěry. Homicidní adaptivní teorie nepředpokládá, že by vražda příslušníka vlastního druhu byla okrajem kontinua, považuje ji za evolučně unikátní robustní strategii ukončující soutěž mezi dvěma jedinci (7, 13, 14).
Lze předpokládat, že homicidní fantazie je projevem tohoto mechanismu, jakmile je cena za vraždu tak vysoká, že odrazuje. Odhaduje se, že na každou skutečnou vraždu připadá mnohonásobně vyšší počet vražd spáchaných jen v pachatelově představě. Při zkoumání 760 vysokoškolských amerických studentů uvedlo alespoň jednu homicidní fantazii 79 % mužů a 58 % žen, 38 % mužů a 18 % žen uvedlo víc než jednu.
Většina mužů prohlásila, že jejich homicidní fantazie trvaly několik minut, 18 % mužů sdělilo, že trvaly několik hodin a déle. Celkem 61 % žen uvedlo jen několikasekundové trvání. Podstatně vyšší podíl mužů než žen (71 % vs. 52 %) uvedlo, že homicidní fantazii spustí pocit osobního ohrožení. Mužské víc než ženské fantazie pobíjely cizí lidi (53 % vs. 33 %), vůdce národa (34 % vs. 17 %), jakož i šéfa (35 % vs. 21 %), ženské fantazie vraždily spolubydlící (34 % vs. 23 % mužských).
Teorie inkluzivní zdatnosti (investuji do příbuzných tím víc, čím jsou mi geneticky bližší) předpovídá větší konflikty s náhradními než biologicky vlastními rodiči. U 44 % lidí, kteří žili s náhradními rodiči, se objevovaly fantazie, že je zabijí. Jestliže s nimi žili déle než 6 let, objevovaly se tyto fantazie u 59 %. Fantazie vraždy biologicky vlastní matky se objevovala ve 31 %, biologicky vlastního otce ve 25 % (18).
Naproti tomu Daly a Wilson (10, 11, 12), rovněž evoluční psychologové, nepovažují zabití druhého člověka za adaptaci, ale za nástroj, jenž je výsledkem přírodního výběru určeným k řešení soutěže mezi muži a sexuální žárlivosti. Z Hamiltonova principu inkluzívní zdatnosti (15; „investuji do příbuzných tím víc, čím jsou mi geneticky bližší a naopak“) plyne, že typy ne-válečného zabíjení jedněch lidí jinými, budou ovlivněny geneticky. Prokázali to jednak na základě historických záznamů z Anglie počínaje 13. stoletím, jednak na základě současných statistik Kanady a USA.
Z anglických historických záznamů plyne, že se na zabíjení často podílel větší počet lidí. Lidé ze skupin, které zabíjely, byli ve vzájemném příbuzenském vztahu 6x častěji, než byly oběti a pachatel. Statistiky z USA a Kanady dokazují, že děti zabíjejí častěji adoptivní nebo náhradní rodiče než rodiče vlastní.
Daly a Wilson (10, 11, 12) užili teorii her a dokázali, že pachatelé i oběti vražd jsou častěji lidé nezaměstnaní, kteří nejsou ženatí nebo vdané: jejich zdroje jsou malé a rizikovým chováním, vyjde-li, mohou mnoho získat.
Agrese nebývá slepý automatismus. Záleží na souvislostech (2). Například domácího násilí na ženách se častěji dopouštějí muži, jejichž hodnota coby partnera je nižší, než je stejná hodnota jejich ženy, případně muže, kteří přišli o zdroj, kterého si žena cení, například o práci. Riziko partnerčiny nevěry roste, agrese má být prevencí.
Většina pachatelů i obětí násilných činů jsou muži, přes jisté kolísání to platí pro všechny zkoumané kultury. Evoluční psychologové to vysvětlují intrasexuální soutěží. U živočišných druhů – lidé mezi ně patří – u nichž samičky „investují“ do potomků podstatně víc než samečkové, jsou samičky cenný a limitující zdroj samčí reprodukce. Ta není omezována schopností přežít, ale mírou přístupu k samičkám. Samečkové těchto druhů včetně mužů jsou schopni zplodit podstatně větší počet potomků než samičky. Výsledkem reprodukčních rozdílů je pohlavní dimorfie – samci jsou kromě jiného statnější než samičky, a to tím víc, čím je polygynie větší. Dalším důsledkem je soutěž o samičky včetně riskantních soubojů.
V současném sociálním prostředí je cena přímé fyzické agrese příliš vysoká, nahrazuje ji proto nepřímá agrese, jejíž přímá cena je nižší (4).
Fylogeneze
Fylogeneze agrese má vztah ke své evoluční funkci. Jakmile je přínos agresivního chování vyšší než riziko, objevuje se agrese u řady živočišných druhů. Agresivní chování se prokazuje u mořských sasanek, mořských polypů i korálů, takže mozek podmínkou agresivního chování není (2).
Fylogeneze agrese odpovídá fylogenezi mozku a sítě tvořící emoce (19, 20). Ty jsou v mozku zvířat a lidí regulovány obvodem, jehož klíčové uzly tvoří
- orbitální, ventromediální a dorzolaterální prefrontální kůra,
- přední cingulární kůra,
- amygdala,
- ventrální striatum/nc. accumbens,
- některé hypotalamické struktury,
- řada jader mozkového kmene, zejména area tegmentalis ventralis Tsai,
- šeď kolem mokovodu, a
- některé oblasti mozečku (obr. 1) (19, 20, 21).
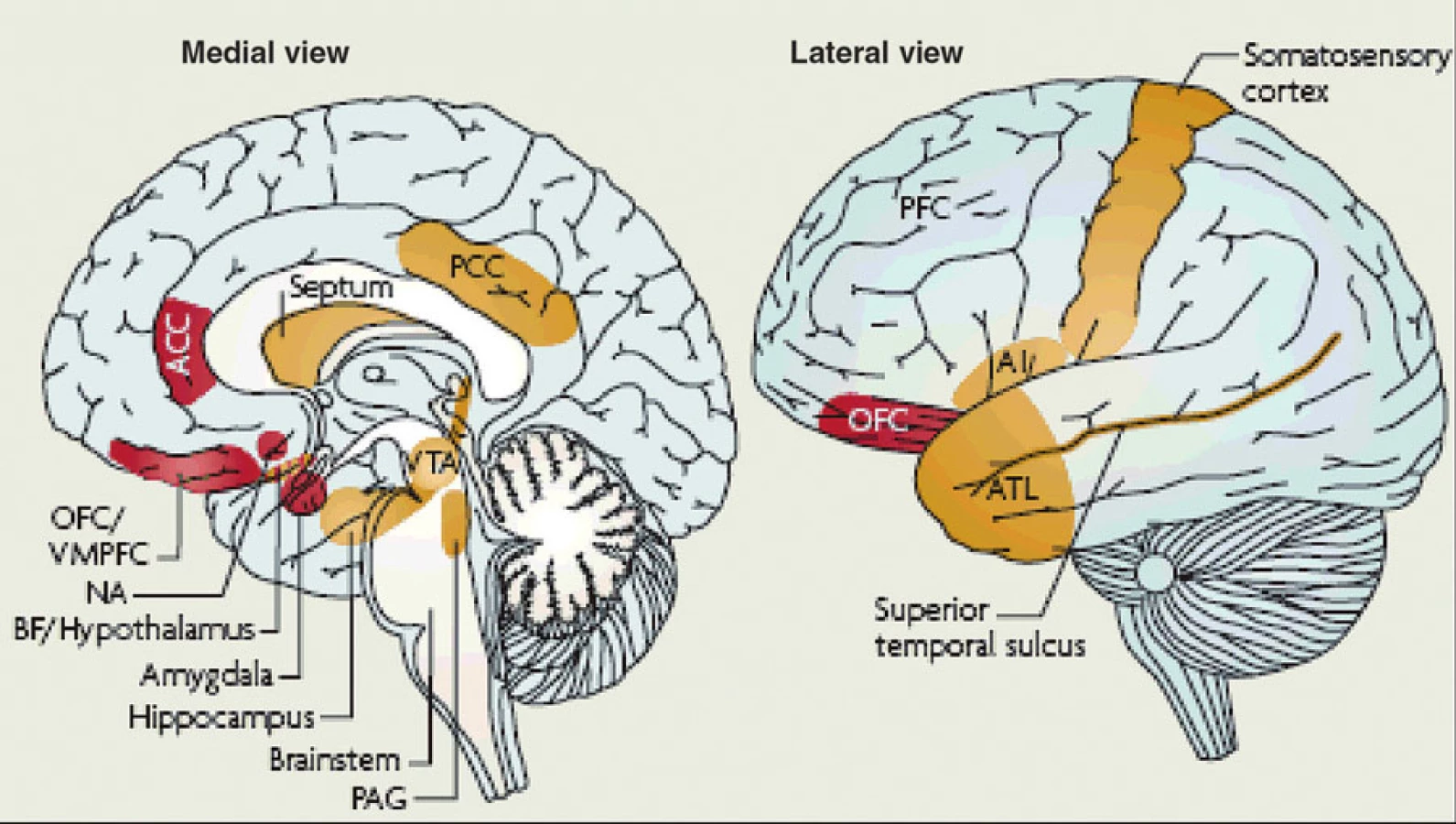
Ovlivnění uzlů této sítě může agresi spustit i zastavit.
Například amygdala řídí činnost podkorových systémů odpovídajících na stres, vyšší nebo nižší míra odpovědi je závislá na kontextu. Orbitofrontální kůra integrující korové a podkorové vstupy kontroluje reaktivní agresi. Důsledkem jejího poškození je impulzivní chování včetně impulzivní agrese (5).
Hermans et al. (17) užili funkční magnetickou rezonanci k měření aktivity ženského mozku, odpovídajícího na pozorování rozzlobené tváře. U žen se aktivovala
- orbitofrontální kůra,
- amygdala,
- hypothalamus,
- šeď kolem mokovodu.
Zvýšení hladiny cirkulujícího testosteronu těchto mladých žen na hladinu blízkou mladým mužům zvyšovalo míru odpovědi podkorových struktur.
Ontogeneze a bezprostřední příčiny
Evoluční perspektiva chápe agresi jako výsledek interakce genů, funkčních systémů mozku a vlivů prostředí. Agresivní chování vůči vrstevníkům se projevuje ve věku 12 měsíců, do 17 měsíce věku roste, v tomto věku už je fyzicky útočných asi 80 % dětí. Míra fyzické agrese je nápadná kolem druhého roku věku, pak ji výchova tlumí, takže mezi 2–11. rokem věku – byť s velkou variabilitou – její míra klesá (9).
Agresivita dětí je ovlivňována typem vazby
- k matce nebo osobě, která ji nahrazuje,
- výchovou,
- vrstevníky, a
- mediálním násilím.
Klíčové pro vývoj násilného chování bývají vztahy v rodině. Násilné chování se v rodinách přenáší z generace do generace. Uplatňuje se jak genotyp, tak fenotyp prarodičovské a rodičovské generace. U dětí záleží na přenosu genů, fetálním prostředí, včetně expozice alkoholu a drog, případných porodních komplikacích a výchovném prostředí, zejména na zanedbávání a zneužívání dětí. Obecně platí, že děti v souvislosti s násilným chováním se před soudem ocitají dvakrát, nejprve jako svědci, poté jako pachatelé (29). Vždy je nutné brát v úvahu genotyp a pohlaví, a to jak rodičů, tak dětí: ne všichni zneužívaní chlapci se v dospělosti chovají násilně, vyšší frekvence je u těch z nich, jejichž exprese genu kódujícího MAO-A je nižší (8).
U dospělých lidí s těžce agresivním a impulzivním chováním se prokazuje snížená míra neurotransmise serotoninu. Otázku, zda je nízká hladina serotoninu mediátorem tohoto chování, nebo zda je reflexí preexistujícího rysu osobnosti, která je dlouholetou debatou, řešila Booijová et al. vyšetřováním dvou skupin dospělých mužů sledovaných 21 let (6). K vyšetřování byla užita pozitronová emisní tomografie. Jednu skupinu tvořili muži, u nichž se v dětství objevovala vysoká míra fyzické agresivity, druhou skupinou byli muži, jejichž míra agresivity v dětství byla nízká. U mužů v první skupině bylo zjištěno významně nižší vychytávání stopující látky v orbitofrontální kůře oboustranně. Stopující látkou byl radioaktivně značený metyl-L-tryptofan. Tito muži sami uváděli vyšší míru impulzivity.
Bez ohledu na tuto neurobiologickou skutečnost se však obě skupiny v dospělosti neodlišovaly
- v hladině plazmatického tryptofanu,
- genotypu,
- míře agrese,
- emoční inteligenci,
- pracovní paměti,
- míře impulzivity určené počítačem,
- psychosociálních funkcích,
- sociálním přizpůsobení,
- výkonu pracovní paměti, a
- osobní ani rodinné anamnéze poruch nálady nebo zneužívání drog.
Z toho plyne, že nízká hladina serotoninu není mediátorem aktuálního chování a lze ji považovat za faktor zranitelnosti, který se může, ale nemusí vyjadřovat. Záleží na dalších biologických faktorech, zkušenosti a vztazích k sociálnímu prostředí v průběhu osobního vývoje.
Kořeny antisociálního chování dospělých nalezl v dětském věku Simonoff et al. (26). Sledoval 225 členů páru dvojčat, která byla vyšetřena v dětství a znovu vyšetřena po 10–25 letech. Prediktorem antisociální poruchy osobnosti a kriminálního chování v rané a střední dospělosti byla ve stejném stupni jednak hyperaktivita, jednak porucha chování dětského věku. Vývojová porucha čtení a nižší IQ bylo prediktorem antisociálního chování v dětství a dospívání.
Počet příznaků hyperaktivity a poruchy chování v dětském věku předpovídal vyústění v dospělosti. Na vyústění má vliv učení, respektive klíčové zkušenosti výchovy, které mohou trajektorii směřující k antisociálnímu chování odvrátit. Omezením této studie je užití retrospektivních dat.
Prasad et al. (25) vyšetřoval kognitivní, motorický a jazykový vývoj 19 batolat a předškoláků, kteří byli ve věku 14–77 měsíců hospitalizováni se známkami tělesného zneužívání, aniž by přitom bylo prokázáno neurologické poškození. Kontrolní skupinu představoval stejný počet stejně starých zdravých dětí. Děti byly vyšetřeny za tři měsíce po propuštění s nemocnice spolu s lidmi, kteří o ně pečovali. U tělesně zneužívaných dětí bylo zjištěno významně nižší skóre v testech jak kognitivních, tak motorických a jazykových funkcí. U dvou dětí ze zneužívané skupiny bylo magnetickou rezonancí prokázáno poškození mozku.
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Zdroje
1. Anderson, C.A., Bushman, B.J. Human aggression. Annu Rev. Psychol. 2002, 53, p. 27-51.
2. Archer, J. The behavioural biology of aggression. Cambridge UK: Cambridge University Press, 1988.
3. Archer, J. The nature of human aggression. Int. J. Law. Psychiatry 2009, 32, p. 202-208.
4. Bjorkvist, K., Osterman, K., Lagerspetz, K.M. Sex differences covert aggression among adults. Aggressive Behavior 1994, 20, p. 27-33.
5. Blair, J.R. The role of orbital frontal cortex in the modulation od antisocial behavior. Brain Cogn. 2004, 55, p. 198-208.
6. Booij, L., Tremblay, R.E., Leyton, M. et al.Brain Serotonin Synthesis in Adult Males Characterized by Physical Aggression during Childhood: A 21-Year Longitudinal Study. PLoS ONE 5(6): e11255. doi:10.1371/journal.pone.0011255 (2010)
7. Buss, D.M., Duntley, J.D. The evolution of aggression. In Schaller M, Kernick DT, Simpson JA (vyd.) Evolution and social Psychology. New York: Psychology Press, 2006, p. 263-285.
8. Caspi, A., McClay, J., Moffitt, T.E. et al. Role of genotype in the cycle of violence in maltreated children. Science 2002, 297, p. 851-854.
9. Coté, S.M., Vaillancourt, T., Leblanc, J.C. et al. The development of physical aggression from toddlerhood to pre-adolescence. A nationwide longitudinal study of Canadian children. J. Abnorm. Child. Psychol. 2006, 34, p. 71-85.
10. Daly, M., Wilson, M. Homicide. New York: Aldine de Gruyter, 1988.
11. Daly, M., Wilson, M. Killing the competition. Female/female and male/male homicide. Hum. Nat. 1990, 1, p. 81-107.
12. Daly, M., Wilson, M. Risk taking, intrasexual competition, and homicide. Nebr. Symp. Motiv. 2001, 47, p. 1-26.
13. Duntley, J.D., Buss, D.M. The evolution of evil. In: The social psychology of good and evil. Miller AG (ed). New York–London: Guilford Press, 2004, p. 102-123.
14. Duntley, J.D., Buss, D.M. The origins of homicide. In J. Duntley and T. Shackelford (vyd.), Evolutionary Forensic Psychology. New York: Oxford University Press 2008, p. 41-64.
15. Hamilton, W.D. The genetical evolution of social behavior I, II. J. Theor. Biol. 1964, 7, p. 1-52.
16. Hare, R.D., Neumann, C.S. Structural models of psychopathy. Curr. Psychiatry Rep. 2005, 7, p. 57-64.
17. Hermans, E.J., Ramsey, N.E., van Honk, J. Exogenous testosterone potentiates response to angry facial expression in the neural circuitry of reactive aggression in humans. Biol. Psychiatry 2008, 63, p. 263-270.
18. Kenrick, D.T., Sheets, V. Homicidal fantasies. Ethol. Sociobiol. 1993;14, p. 231-246.
19. Koukolík, F. Neuronální koreláty agrese a násilného chování. In.: Sociální mozek. Praha: Karolinum, 2006, s. 243-262.
20. Koukolík, F. Evoluce a evoluční teorie pro lékaře. VIII. Evoluce lidského mozku. Prakt. lék. 2010, 90, 8, s. 451-454.
21. Koukolík, F. Funkční systémy lidského mozku. Galén, Praha 2011 (v tisku), kapitola Emoce.
22. Maynard-Smith, J. Evolution and the theory of games. Cambridge and New York: Cambridge University Press, 1982.
23. Moyer, K.E. The psychobiology of aggression. New York: Harper and Row Publishers, 1976.
24. Pessoa, L. On the relationship between cognition and emotion. Nat. Rev. Neurosci. 2008, 9, p. 148-158.
25. Prasad, M.R., Kramer, L.A., Ewing-Cobbs, L. Cognitive and neuroimaging findings in physically abused preschoolers. Arch. Dis. Child. 2005, 90, p. 82-85.
26. Simonoff, E., Elander, J., Holmshaw, J. et al. Predictors of antisocial personality. Continuities from childhood to adult life. Brit. J. Psychiatry 2004, 184, p. 118-127.
27. Tinbergen, N. On the aims and methods of ethology. Z Tierpsychol. 1963, 20, p. 410-433.
28. Victoroff, J. Aggression, science, and law: the origins framework. Int. J. Law Psychiatry. 2009, 32, p. 189-197.
29. Volavka, J. The neurobiology of violence: an update. J. Neuropsychiatry Clin. Neurosci. 1999; 11, p. 307-313.
30. Wilson, E.O. Sociobiology. Cambridge: Harvard University Press, 1975.
31. WHO: World report on violence and health. WHO, Geneva 2002
Štítky
Praktické lekárstvo pre deti a dorast Praktické lekárstvo pre dospelýchČlánok vyšiel v časopise
Praktický lékař

2011 Číslo 3
- Metamizol jako analgetikum první volby: kdy, pro koho, jak a proč?
- Fixní kombinace paracetamol/kodein nabízí synergické analgetické účinky
- Tramadol a paracetamol v tlumení poextrakční bolesti
- Realita liečby bolesti v paliatívnej starostlivosti v Nemecku
Najčítanejšie v tomto čísle
- Netuberkulózní mykobakteriózy
- Začátky organizovaného boje proti tuberkulóze v Čechách
- Případ krční lymfadenopatie
- Současné trendy péče o dítě s orofaciálním rozštěpem