
Nové metody studia metylace DNA – MS-HRM analýza a elektrochemie
Novel Approaches in DNA Methylation Studies – MS-HRM Analysis and Electrochemistry
Cytosine methylation in DNA is an epigenetic mechanism regulating gene expression and plays a vital role in cell differentiation or proliferation. Tumor cells often exhibit aberrant DNA methylation, e.g. hypermethylation of tumor suppressor gene promoters. New methods, capable of determining methylation status of specific DNA sequences, are thus being developed. Among them, MS-HRM (methylation-specific high resolution melting) and electrochemistry offer relatively inexpensive instrumentation, fast assay times and possibility of screening multiple samples/DNA regions simultaneously. MS-HRM is due to its sensitivity and simplicity an interesting alternative to already established techniques, including methylation-specific PCR or bisulfite sequencing. Electrochemistry, when combined with suitable electroactive labels and electrode surfaces, has been applied in several unique strategies for discrimination of cytosines and methylcytosines. Both techniques were successfully tested in analysis of DNA methylation within promoters of important tumor suppressor genes and could thus help in achieving more precise diagnostics and prognostics of cancer. Aberrant methylation of promoters has already been described in hundreds of genes associated with tumorigenesis and could serve as important biomarker if new methods applicable into clinical practice are sufficiently advanced.
Key words:
DNA methylation – 5-methylcytosine – HRM analysis – melting temperature – DNA duplex – electrochemistry – nucleic acid hybridization
This work was supported by MEYS – NPS I – LO1413.
The authors declare they have no potential conflicts of interest concerning drugs, products, or services used in the study.
The Editorial Board declares that the manuscript met the ICMJE recommendation for biomedical papers.
Submitted:
6. 5. 2016
Accepted:
16. 5. 2016
Authors:
M. Bartošík; E. Ondroušková
Authors‘ workplace:
Regionální centrum aplikované molekulární onkologie, Masarykův onkologický ústav, Brno
Published in:
Klin Onkol 2016; 29(Supplementum 4): 64-71
Category:
Review
doi:
https://doi.org/10.14735/amko20164S64
Overview
Metylace cytozinů v DNA je jedním z epigenetických mechanizmů regulujících expresi genů, a hraje tak důležitou roli v diferenciaci nebo proliferaci buněk. V nádorových buňkách často dochází ke změnám v metylaci DNA, kupříkladu nadměrnou metylací (hypermetylací) promotorů tumor supresorových genů. Proto jsou vyvíjeny nové metody analýzy, které by byly schopny určit rozsah metylace konkrétní sekvence DNA. K těmto metodám se řadí i relativně levné a rychlé techniky MS-HRM (methylation-specific high resolution melting) a elektrochemie. Ve srovnání s jinými používanými metodami jsou vhodné pro screening většího počtu vzorků či více cílových oblastí DNA. MS-HRM svou citlivostí a relativní nenáročností konkuruje ostatním zavedeným metodám, jako jsou metylačně-specifická PCR nebo přímá bisulfitová sekvenace. Elektrochemie nabízí nenáročné přístrojové vybavení a při použití vhodných elektroaktivních molekul a elektrodových povrchů i několik zajímavých strategií rozlišení cytozinů a metylcytozinů. Obě techniky byly již úspěšně použity při stanovení metylace DNA promotorů důležitých tumor supresorových genů a mohly by tak v budoucnu přispět ke zpřesnění diagnostiky a prognostiky onkologických onemocnění. Aberantní metylace promotorů byla popsána již u stovek genů se vztahem k nádorovým onemocněním a s rozvojem metod, jež by umožňovaly jejich detekci i v klinické praxi, by se řada z nich mohla stát novými důležitými biomarkery.
Klíčová slova:
DNA metylace – 5-metylcytozin – HRM analýza – teplota tání – DNA duplex – elektrochemie – hybridizace nukleových kyselin
Úvod
Epigenetické mechanizmy hrají klíčovou roli v normálním vývoji jedince i při vzniku různých onemocnění. Regulují expresi genů reverzibilní kovalentní modifikací DNA, RNA nebo proteinů (bez změny pořadí nukleotidů anebo aminokyselin), které jsou ovšem dědičné, a ovlivňují tak fenotyp, aniž by měnily genotyp. Epigenetické mechanizmy se uplatňují na celé řadě úrovní (před transkripcí, post-transkripční i post-translační), a jsou tak důležité např. v morfogenezi, při diferenciaci buněk anebo při jejich proliferaci. Mezi epigenetické mechanizmy se řadí např. metylace DNA, acetylace histonů nebo fosforylace, sumoylace či ubikvitinace různých proteinů.
V případě metylace DNA zde máme na mysli reakci katalyzovanou DNA metyltransferázou (DNMT), která přenáší metylovou skupinu (-CH3) ze substrátu S-adenozyl-methioninu (SAM) na uhlík cytozinu v pozici 5’, nacházejícího se v dinukleotidu CpG, za vzniku 5-metylcytozinu (obr. 1). Metylace cytozinů v DNA „tlumí“ expresi genů, přičemž se předpokládají dva hlavní mechanizmy – přímá interference vazby transkripčních faktorů na DNA a aktivace histonových deacetyláz (které snižují expresi genů ovlivněním kompaktnosti DNA). Distribuce CpG dinukleotidů je v rámci genomu značně asymetrická, jsou obvykle seskupeny do krátkých úseků, tzv. CpG „ostrůvků“, nacházejících se nejčastěji v promotorech genů. Ve většině případů jsou tyto cytoziny nemetylované, výjimky tvoří plně metylované ostrůvky např. v některých genech u ženských chromozomů X nebo u imprintovaných genů na jedné parentální alele [1].

Abnormální metylace DNA byla detekována i v nádorových buňkách. Ty paradoxně vykazují jednak zvýšenou metylaci (hypermetylaci) promotorů tumor supresorových genů [1] blokující expresi proteinů důležitých např. při opravě poškození DNA nebo apoptóze, na druhé straně však i globálně sníženou metylaci (hypometylaci) genomu vedoucí k vyšší expresi původně utlumených, potenciálně nebezpečných genů (např. virových nebo imprintovaných genů, repetitivních sekvencí atd.) [2], popřípadě k chromozomální destabilizaci [3]. Hypermetylace promotorů byla prokázána např. v genu kódujícím protein p16 u různých typů nádorů (karcinom mléčné žlázy, žaludku, plic atd.) [4], BRCA1 (karcinom mléčné žlázy a vaječníků) [5], v genu kódujícím O6-metylguanin DNA metyltransferázu (MGMT) v glioblastomech [6], glutation-S-transferázu [7] nebo VHL (karcinom ledvin) [8]. Objevují se důkazy, že metylace DNA ovlivňuje i expresi mikroRNA, důležitých posttranskripčních regulátorů genové exprese [9].
Globální hypometylace DNA byla naopak nalezena např. u nádorů prostaty, hepatocelulárních karcinomů nebo karcinomu děložního hrdla [10]. Snížená metylace nebo až demetylace může postihnout i promotory onkogenů. Naše skupina prokázala, že demetylace onkogenu E6 v genomu lidského papilomaviru 16 (HPV-16) způsobujícího rakovinu děložního hrdla vede k jeho zvýšené expresi, a může tak sloužit jako prediktivní biomarker transformace prekancerózních lézí na karcinom [11]. Analýza metylace DNA proto skýtá obrovský potenciál využitelný např. pro včasnou diagnostiku nádorů a případné určení jejich invazivity, progrese, metastatického potenciálu nebo chemosenzitivity [12].
Současné metody studia metylace DNA
Existují metody, které sice dokážou kvantifikovat globální metylaci DNA, např. HPLC [13], ovšem neposkytují informace o konkrétních metylovaných pozicích v rámci genomu, a nejsou tak vhodné pro funkční analýzy. Budeme se proto věnovat pouze těm přístupům, které umožňují určit míru metylace konkrétního místa v DNA, tj. obvykle sekvence genu nebo jeho promotoru. Velká většina z těchto metod je založena na 1. použití metyl senzitivních restrikčních endonukleáz, 2. reakci DNA s hydrogensiřičitanem sodným (tzv. bisulfitové konverzi) anebo 3. chromatinové imunoprecipitaci s využitím protilátek vůči metylcytozinu. Všechny tyto přístupy byly vyvinuty v 70. a 80. letech 20. století a od té doby zaznamenaly značný rozvoj.
Restrikční analýza
Metyl senzitivní restrikční endonukleázy (restriktázy) jsou speciální enzymy, které rozeznávají a štěpí konkrétní restrikční místa v DNA v závislosti na přítomnosti metylcytozinu. Obvykle jde o enzymy štěpící pouze nemetylovanou DNA, např. HpaII (rozeznávající a štěpící sekvenci CCGG), BstUI (CGCG) nebo HhaI (GCGC), přičemž přítomnost metylcytozinu v rozeznávané sekvenci štěpení DNA blokuje. Naproti tomu existují i endonukleázy, které naopak štěpí pouze metylovanou DNA, např. McrBC rozeznávající a štěpící dvě (G/A)mC místa vzdálená od sebe až 3 kb [11,14]. Zřejmou nevýhodou restrikčních analýz je nutná přítomnost daného restrikčního místa v analyzovaném genu nebo promotoru. Původní časově i materiálně náročnější Southernovu analýzu štěpených fragmentů [15] nahradila restrikční analýza v kombinaci s kvantitativní PCR (qPCR). DNA je nejdřív štěpena vhodnou endonukleázou a následně pouze nefragmentovaná DNA (tudíž pouze metylovaná anebo nemetylovaná DNA, v závislosti na použité restriktáze) je amplifikována a současně kvantifikována pomocí qPCR [16].
Další metodou je tzv. MS-MLPA (methylation-sensitive multiplex ligation-dependent probe amplification) čili kombinace restrikce, ligace a amplifikace [17]. MLPA test je založen na aplikaci dvou oligonukleotidů, které hybridizují k přilehlým cílovým sekvencím ve zkoumané DNA a pomocí ligázy jsou spojeny do jedné sondy. Současně se přidává i restriktáza, která štěpí nemetylovanou ligovanou DNA. V průběhu následné PCR reakce jsou amplifikovány pouze neštěpené sondy (obsahující primer sekvence), které hybridizovaly s metylovanou DNA. Metoda je sice semi-kvantitativní, ale umožňuje analýzu až 60 různých fragmentů DNA v jediné multiplexní PCR reakci pomocí následné kapilární elektroforézy, a to na základě různých délek oligonukleotidů (obr. 2).

Bisulfitová konverze
Hydrogensiřičitan sodný (sodium bisulfite) deaminuje v sérii kroků cytozin na uracil, zatímco metylcytozin je chemicky chráněn a zůstává nezměněn. Takto modifikovaná DNA je následně použita jako templát pro PCR reakci, přičemž uracil je amplifikován jako tymin a metylcytozin jako cytozin (obr. 3). Tím jsou získány dvě různé sekvence DNA v závislosti na původní metylaci. Tato specifická konverze je základem mnoha technik analýzy DNA metylace.
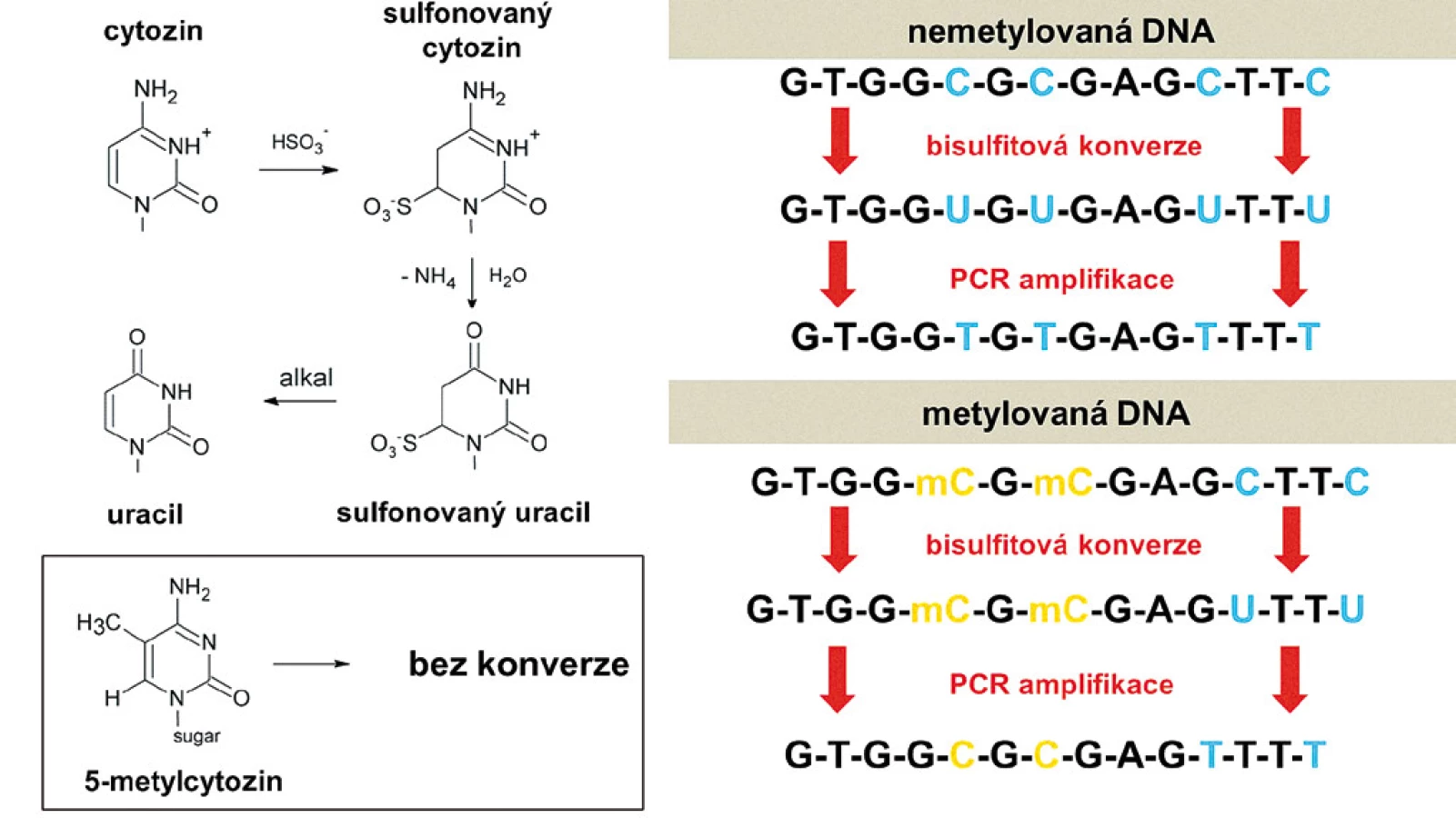
Mezi nejpoužívanější patří metylačně-specifická PCR (MSP) [18], která amplifikuje bisulfitem konvertovanou DNA pomocí metyl-specifických primerů, přičemž tyto primery jsou navrženy zvlášť pro konvertovanou nemetylovanou DNA (která obsahuje uracily, a proto má primer více adeninů) a zvlášť pro konvertovanou metylovanou DNA (primer obsahuje více guaninů). Amplikony jsou následně vizualizovány pomocí gelové elektroforézy. Jedná se o rychlou a citlivou metodu bez nutnosti speciálního příslušenství, není však dostatečně kvantitativní. Tou je naopak metoda MethyLight [19] kombinující MSP (tj. konverzi bisulfitem s následnou amplifikací pomocí metyl-specifických primerů) a fluorescenční detekci pomocí Taqman sondy (DNA oligonukleotid značený na jednom konci fluoroforem a na druhém konci zhášedlem). Během PCR Taqman sonda hybridizuje s templátovou DNA v rámci amplifikované sekvence, přičemž DNA polymeráza během replikace sondu rozštěpí, fluorofor je tím od zhášedla oddělen, což vede k nárůstu fluorescence. I když se jedná o metodu dražší, Taqman sondy zabezpečují vyšší specificitu amplifikace.
Bisulfitové pyrosekvenování poskytuje relativně přesná data o metylaci jednotlivých cytozinů, i když pouze u krátkých sekvencí do délky cca 250 bází. Po bisulfitové konverzi DNA následuje PCR, u které je jeden z primerů biotinylován, a může tak být imobilizován na streptavidinových kuličkách. Do reakční směsi jsou postupně přidávány jednotlivé deoxynukleotidtrifosfáty (dATP, dCTP, dGTP, dTTP), přičemž v reakční směsi je v daný okamžik přítomen pouze jeden z těchto nukleotidů. Po inkorporaci komplementárního nukleotidu se uvolní pyrofosfát (PPi), který je enzymem ATP sulfurylázou konvertován na ATP. ATP je následně využito luciferázou k oxidaci luciferinu na oxyluciferin, čímž dochází k uvolnění světla, které je snímáno kamerou. Nespotřebované nukleotidy jsou na závěr degradovány apyrázou a z kuliček odmyty, aby do reakční směsi mohly vstoupit jiné nukleotidy [20].
Kombinaci bisulfitové konverze s restrikčními endonukleázami představuje tzv. COBRA analýza (combined bisulfite restriction analysis). Po bisulfitové konverzi a PCR amplifikaci jsou amplikony štěpeny restriktázou, která specificky štěpí restrikční místo ovlivněné konverzí. Elektroforézou v denaturujícím gelu lze z poměru štěpených a neštěpených PCR produktů zjistit stupeň metylace [21].
Proteiny vázající metylcytozin
Metylcytoziny v DNA mohou být specificky zachyceny pomocí protilátek vůči metylcytozinu, popřípadě s využitím proteinů vázajících metylovou skupinu (methyl-binding proteins). V obou případech se v podstatě jedná o obohacení vzorku o metylovanou DNA a její následnou analýzu pomocí různých metod. Imunoprecipitace metylované DNA (methylated DNA immunoprecipitation – MeDIP) představuje metodu globální analýzy metylace. DNA je fragmentována ultrazvukem, po denaturaci inkubována s protilátkou proti metylcytozinu a následně se sekundární protilátkou navázanou na magnetickou kuličku. Metylovaná DNA je tak zachycena a nemetylovaná DNA je následně odmyta. Získaná DNA je sekvenována nebo analyzována pomocí DNA microarrays [22].
HRM analýza
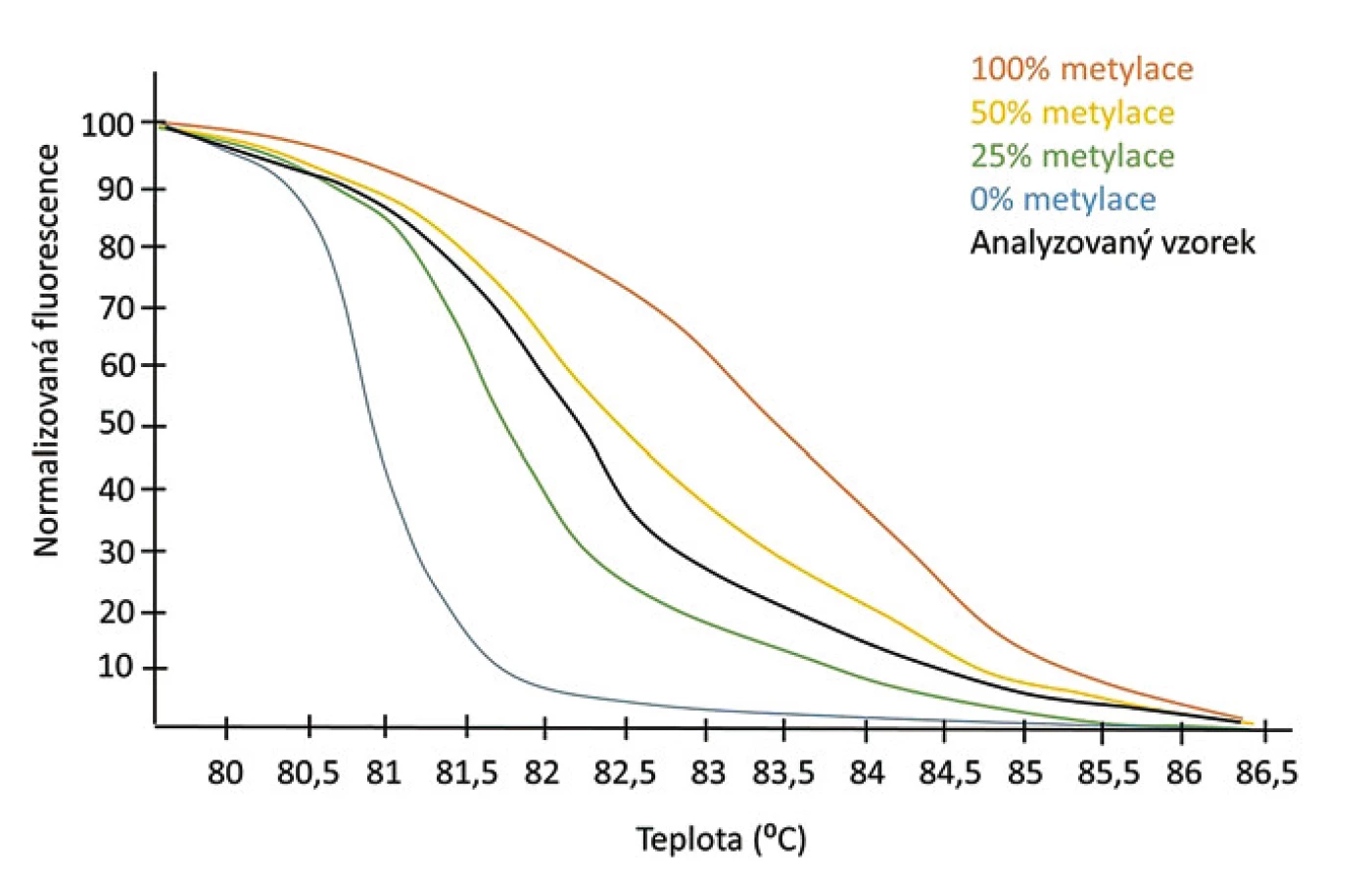
Jednou z poměrně citlivých a nepříliš nákladných metod analýzy metylace DNA je MS-HRM (methylation-specific high resolution melting) využívající rozdílných teplot tání dvouřetězcových DNA v závislosti na jejich sekvenci. Podíl párů cytozin-guanin (C-G) a tymin-adenin (T-A) obecně určuje pevnost vazby mezi dvěma řetězci DNA. Pár C-G je spojen třemi vodíkovými vazbami, zatímco T-A jen dvěma. Čím vyšší je tedy podíl C-G párů v analyzovaném řetězci, tím je vazba dvouřetězce silnější a tím vyšší je teplota tání, Tm, které je potřeba dosáhnout, aby dvouřetězcová DNA disociovala na jednotlivé řetězce. Pokud je k detekci DNA použito fluorescenční interkalační barvivo, pak je možné srovnat Tm jednotlivých dvouřetězců stanovením teploty, při které dochází k poklesu fluorescence danému rozpadem dvouřetězce a uvolněním interkalačního barviva.
Vývoj interkalačních barviv, která neinhibují průběh PCR, a přístrojů s dostatečnou citlivostí zachycujících i jemné změny fluorescence vedl k využití metody HRM pro detekci jednonukleotidových polymorfizmů v genotypizačních studiích [23,24]. Stejný princip lze využít i pro rozlišení mezi metylovanou a nemetylovanou alelou, přičemž MS-HRM poprvé využili k detekci metylace MGMT promotoru Wojdacz et al [25]. V této variantě metody HRM je DNA nejprve podrobena bisulfitové konverzi a poté je v přítomnosti fluorescenčního interkalačního barviva amplifikován analyzovaný úsek. Po ukončení amplifikace je sledován pokles fluorescence jednotlivých vzorků při zvyšující se teplotě. Ke stanovení procenta metylace vzorku je vždy jako standard použita plně metylovaná a plně demetylovaná DNA, které jsou smíchány v určitých poměrech a od nichž lze stupeň metylace vzorku odečíst (obr. 4). Výhodou MS-HRM jsou poměrně nízké náklady a dostatečná citlivost pro detekci 0,1% metylace templátu. Nevýhodou je, že touto metodou lze zjistit pouze procento metylace dané sekvence, ale nikoliv, které konkrétní CpG dinukleotidy jsou metylované a které ne. Dále, stejně jako u ostatních metod založených na PCR dochází k nerovnoměrné efektivitě amplifikace metylované a nemetylované DNA sekvence, kdy nemetylovaná DNA je obecně amplifikována s vyšší účinností. Tento problém je řešitelný upraveným designem primerů tak, aby obsahovaly jeden až dva CpG dinukleotidy u 5’-konce, čímž se stanou o něco specifičtější pro metylovanou alelu. Takto navržené primery spolu s optimalizací teploty renaturace pak umožní vyrovnání účinnosti PCR u metylované a nemetylované DNA [26]. MS-HRM lze nejlépe využít pro kvantifikaci metylace u DNA sekvencí bohatých na CpG dinukleotidy, které tvoří pouze homoduplexy, a mohou tak být srovnány s plně metylovanou/demetylovanou DNA. V případě sekvencí s nízkým obsahem CpG (méně než tři) se mohou tvořit i heteroduplexy a křivky tání jsou potom komplexnější a obtížněji interpretovatelné. Přesnější stanovení počtu metylovaných/nemetylovaných alel i ve vzorku s nízkým obsahem CpG umožňuje modifikace MS-HRM využívající digitální PCR (dMS-HRM), při níž je vzorek před amplifikací naředěn tak, že je následně amplifikována vždy jen jedna molekula DNA. Tím se zároveň vyřeší i nerovnoměrná efektivita amplifikace metylovaných vs. nemetylovaných DNA sekvencí [27].
Od dob těchto pilotních studií se využití metodiky MS-HRM postupně rozšiřuje a přibývá prací, ve kterých byla úspěšně aplikována na detekci metylace promotorů genů spojených většinou s nádorovými onemocněními. Konkrétně byla takto popsána metylace promotoru genu AKAP12 u karcinomu prostaty [28], u pacientek s karcinomem prsu pak byla touto metodou detekována např. metylace promotoru genu CST6 [29] nebo SOX17 [30]. Yang et al provedli MS-HRM analýzu metylace promotorů několika genů s funkcí nádorových supresorů, přičemž získali zajímavé výsledky, které by mohly být využity při včasné detekci karcinomů nosohltanu [31]. Citlivost metody potvrzuje i její využití u pacientů s kolorektálním karcinomem při analýze metylací cirkulující nádorové DNA v periferní krvi [32] nebo ze stolice [33]. U pacientek s rakovinou děložního hrdla pak byla využita k detekci metylovaného virového genu L1 u HPV-16, čímž by mohla přispět k přesnější prognostice tohoto onemocnění [34,35]. dMS-HRM byla využita při detailní analýze metylace promotoru genu kódujícího BRCA1 nebo CDKN2B [27,36].
Elektrochemie
Elektrochemie se na rozdíl od výše popsaných optických metod zabývá přenosem elektronů mezi studovanou látkou a elektrodou, a to v prostředí elektrolytu (tj. vodivého roztoku). Studovaná látka může elektrodě elektrony odevzdávat (oxidovat se), nebo od elektrody elektrony přijímat (redukovat se), přičemž se tento přenos elektronů projeví změnou sledovaného signálu (obr. 5). Výhodou elektrochemie je jednoduchá a relativně levná instrumentace, rychlé měření a možnost miniaturizace systému s paralelní detekcí vzorků. I proto je čím dál častěji aplikována v oblasti molekulární onkologie, kde se obvykle používá při detekci různých biomarkerů (proteinů [37], DNA [38,39] nebo mikroRNA [40]), studiu interakcí DNA s proteiny [41] a poškození DNA nízkomolekulárními látkami [38], ale i při testování potenciálních protinádorových léčiv [42].

Bylo rovněž publikováno několik strategií využívajících elektrochemickou analýzu metylace DNA. Nejjednodušší je přímé rozlišení cytozinu a metylcytozinu na povrchu určitých elektrod na bázi uhlíku, a to díky rozdílnému potenciálu jejich oxidace [43]. Výsledná voltametrická křivka (křivka závislosti proudu na potenciálu) pak obsahuje dva oddělené píky, jeden od cytozinu a druhý od metylcytozinu. Tento přístup, který jako jeden z mála nevyžaduje hybridizaci s komplementární DNA, ovšem vyžaduje vysoké vstupní koncentrace DNA a neumožňuje lokalizaci CpG dinukleotidů v rámci analyzované sekvence.
Velice častou strategií je elektrochemické sledování aktivity a inhibice metyltransferáz [44–50]. Povrch elektrody, popřípadě magnetických kuliček je modifikován nemetylovaným duplexem DNA obsahujícím restrikční místo pro vybranou restriktázu, přičemž jedno z vláken DNA je obvykle elektrochemicky značeno. V přítomnosti DNMT nastává metylace cytozinů v CpG místech a následná inkubace s restriktázou vede k tomu, že nemetylovaný duplex je štěpen, zatímco metylovaný ne. Promytím elektrody/kuliček je odstraněna značená část nemetylované DNA, což vede v porovnání s metylovaným vzorkem k poklesu signálu. Jednotlivé práce se pak liší typem studovaného genu, zvolené restriktázy nebo použité elektrochemické značky. Skupina J. Bartonové aplikovala popsanou strategii pro studium hyperaktivity DNMT1 přímo v buněčných lyzátech kolorektálního karcinomu, přičemž zjistili výrazně vyšší aktivitu DNMT1 v nádorové tkáni oproti přilehlé zdravé tkáni [45]. qPCR ani Western blotting analýza ovšem tuto hyperaktivitu nepotvrdily, což autoři vysvětlují vyšší senzitivitou elektrochemické metody.
eMethylsorb je nedávno navržený test pro rozlišení metylované a nemetylované DNA na základě jejich rozdílných adsorpčních vlastností. Obě DNA jsou modifikovány bisulfitem a následně amplifikovány asymetrickou PCR, která vygeneruje jednořetězcové amplikony obohacené buď o guanin (pro metylovanou DNA), anebo adenin (pro nemetylovanou DNA). Rozdílné afinity těchto amplikonů ke zlatému povrchu elektrody vedou k větší adsorpci „adeninových“ amplikonů (od nemetylované DNA), a tudíž k větší elektrostatické repulzi se záporně nabitou elektroaktivní značkou (tj. k poklesu signálu). Prozatím byla metoda testována na DNA z nádorových buněk MCF-7, u které rozlišila 0%, 10%, 25%, 50%, 75% a 100% stupeň metylace [51,52].
Bisulfitová konverze byla použita i pro analýzu metylace genu TP53 [53], a to bez použití PCR amplifikace. Autoři nepoužili DNA sondu, ale její strukturní analog složený z peptidové kostry a DNA nukleotidů, tzv. PNA (peptide nucleic acid) sondu. Tato PNA sonda byla komplementární k metylované DNA po konverzi bisulfitem (naopak nemetylovaná DNA se po konverzi lišila ve dvou nukleotidech, a tudíž nebyla plně komplementární k PNA sondě). Následně aplikovali kladně nabitou elektroaktivní značku, která se elektrostaticky vázala na záporně nabitou metylovanou DNA daleko více než na nemetylovanou DNA (ta s PNA sondou totiž hybridizovala v mnohem menší míře). Výhodou PNA sondy je její elektroneutralita – proto se na ni kladně nabitá značka nevázala, což vedlo ke snížení falešně negativních výsledků. Metoda ovšem nebyla testována na biologických vzorcích, a proto zůstává otázkou, jestli bude efektivní nejenom u nádorových linií, ale zejména v klinickém materiálu od pacientů.
Naše laboratoř rovněž aplikovala bisulfitovou konverzi bez nutnosti použití PCR [54]. Metoda spočívá v tom, že metylcytozin je na rtuťových elektrodách redukovatelný, zatímco uracil po bisulfitové konverzi nikoliv. Metylovaná DNA tak po konverzi poskytovala vyšší signál než nemetylovaná DNA, přičemž množství metylcytozinů v DNA korelovalo s velikostí signálu. I tuto metodu je potřeba optimalizovat pro její použití v biologických vzorcích.
Závěr a výhled do budoucnosti
Studium metylace DNA jakožto epigenetické modifikace regulující genovou expresi je nejen zajímavou, ale i nesmírně důležitou oblastí současného onkologického výzkumu. Čím dál tím více vědeckých prací potvrzuje vliv abnormální metylace na expresi důležitých nádorových supresorů nebo onkoproteinů, a vývoj nových metod pro analýzu metylace DNA tak může vést k efektivnější diagnostice nádorových onemocnění anebo k lepší predikci odpovědí na jejich léčbu. Prozatím však chybí dostatečně ověřené testy použitelné v klinické praxi. Jedním z mála je Epi proColon – diagnostický krevní test nedávno schválený americkým Úřadem pro kontrolu potravin a léčiv pro detekci metylace promotoru genu pro Septin 9 u pacientů s kolorektálním karcinomem [55].
Protože důležitým faktorem těchto testů bude i jejich finální cena, vyvíjejí se i metody relativně časově a laboratorně nenáročné, a tudíž celkově levnější. Mezi tyto metody patří jednak MS-HRM, sledující rozdílné teploty tání duplexů DNA v závislosti na metylaci konkrétních CpG míst, a různé elektrochemické techniky. I navzdory značnému pokroku ve vývoji těchto metod však bude potřeba jejich přísná validace v klinickém materiálu. Dá se očekávat, že ruku v ruce s dalším pokrokem ve výzkumu metylace DNA a jejího vlivu na karcinogenezi dojde k dalšímu rozvoji a využití těchto metod v diagnostice a prognostice onkologických onemocnění.
Práce byla podpořena grantem MŠMT – NPU I – LO1413.
Autoři deklarují, že v souvislosti s předmětem studie nemají žádné komerční zájmy.
Redakční rada potvrzuje, že rukopis práce splnil ICMJE kritéria pro publikace zasílané do biomedicínských časopisů.
Mgr. Martin Bartošík, Ph.D.
Regionální centrum aplikované molekulární onkologie
Masarykův onkologický ústav
Žlutý kopec 7
656 53 Brno
e-mail: martin.bartosik@mou.cz
Obdrženo: 6. 5. 2016
Přijato: 16. 5. 2016
Sources
1. Herman JG, Baylin SB. Mechanisms of disease: gene silencing in cancer in association with promoter hypermethylation. New Engl J Med 2003; 349 (21): 2042–2054.
2. Walsh CP, Chaillet JR, Bestor TH. Transcription of IAP endogenous retroviruses is constrained by cytosine methylation. Nat Genet 1998; 20 (2): 116–117.
3. Xu GL, Bestor TH, Bourchis D et al. Chromosome instability and immunodeficiency syndrome caused by mutations in a DNA methyltransferase gene. Nature 1999; 402 (6758): 187–191.
4. Belinsky SA, Nikula KJ, Palmisano WA et al. Aberrant methylation of p16 (INK4a) is an early event in lung cancer and a potential biomarker for early diagnosis. Proc Natl Acad Sci U S A 1998; 95 (20): 11891–11896.
5. Esteller M, Silva JM, Dominguez G et al. Promoter hypermethylation and BRCA1 inactivation in sporadic breast and ovarian tumors. J Natl Cancer Inst 2000; 92 (7): 564–569.
6. Hegi ME, Diserens A, Gorlia T et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. New Engl J Med 2005; 352 (10): 997–1003.
7. Esteller M, Corn PG, Urena JM et al. Inactivation of glutathione S-transferase P1 gene by promoter hypermethylation in human neoplasia. Cancer Res 1998; 58 (20): 4515–4518.
8. Sato Y, Yoshizato T, Shiraishi Y et al. Integrated molecular analysis of clear-cell renal cell carcinoma. Nat Genet 2013; 45 (8): 860–867. doi: 10.1038/ng.2699.
9. Suzuki H, Maruyama R, Yamamoto E et al. DNA methylation and microRNA dysregulation in cancer. Molecular Oncology 2012; 6 (6): 567–578. doi: 10.1016/j.molonc.2012.07.007.
10. Ehrlich M. DNA methylation in cancer: too much, but also too little. Oncogene 2002; 21 (35): 5400–5413.
11. Hublarova P, Hrstka R, Rotterova P et al. Prediction of human papillomavirus 16 E6 gene expression and cervical intraepithelial neoplasia progression by methylation status. Int J Gynecol Cancer 2009; 19 (3): 321–325. doi: 10.1111/IGC.0b013e31819d8a5c.
12. Delpu Y, Cordelier P, Cho WC et al. DNA methylation and cancer diagnosis. Int J Mol Sci 2013; 14 (7): 15029–15058. doi: 10.3390/ijms140715029.
13. Ehrlich M, Gamasosa MA, Huang LH et al. Amount and distribution of 5-methylcytosine in human DNA from different types of tissues or cells. Nucleic Acids Res 1982; 10 (8): 2709–2721.
14. Badal V, Chuang LS, Tan EH et al. CpG methylation of human papillomavirus type 16 DNA in cervical cancer cell lines and in clinical specimens: genomic hypomethylation correlates with carcinogenic progression. J Virol 2003; 77 (11): 6227–6234.
15. Li E, Bestor TH, Jaenisch R. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell 1992; 69 (6): 915–926.
16. Bruce S, Hannula-Jouppi K, Lindgren CM et al. Restriction site-specific methylation studies of imprinted genes with quantitative real-time PCR. Clin Chem 2008; 54 (3): 491–499. doi: 10.1373/clinchem.2007.098491.
17. von Känel T, Huber AR. DNA methylation analysis. Swiss Med Wkly 2013; 143: w13799. doi: 10.4414/smw.2013. 13799.
18. Herman JG, Graff JR, Myohanen S et al. Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci U S A 1996; 93 (18): 9821–9826.
19. Eads CA, Danenberg KD, Kawakami K et al. Methy Light: a high-throughput assay to measure DNA methylation. Nucleic Acids Res 2000; 28 (8): e32.
20. Tost J, Gut IG. DNA methylation analysis by pyrosequencing. Nat Protoc 2007; 2 (9): 2265–2275.
21. Xiong ZG, Laird PW. COBRA: a sensitive and quantitative DNA methylation assay. Nucleic Acids Res 1997; 25 (12): 2532–2534.
22. Jacinto FV, Ballestar E, Esteller M. Methyl-DNA immunoprecipitation (MeDIP): hunting down the DNA methylome. Biotechniques 2008; 44 (1): 35–43.
23. Wittwer CT, Reed GH, Gundry CN et al. High-resolution genotyping by amplicon melting analysis using LCGreen. Clin Chem 2003; 49 (6): 853–860.
24. Montgomery JL, Sanford LN, Wittwer CT. High-resolution DNA melting analysis in clinical research and diag - nostics. Expert Rev Mol Diagn 2010; 10 (2): 219–240. doi: 10.1586/erm.09.84.
25. Wojdacz TK, Dobrovic A. Methylation-sensitive high resolution melting (MS-HRM): a new approach for sensitive and high-throughput assessment of methylation. Nucleic Acids Res 2007; 35 (6): E41.
26. Wojdacz TK, Hansen LL, Dobrovic A. A new approach to primer design for the control of PCR bias in methylation studies. BMC Res Notes 2008; 1 (1): 54. doi: 10.1186/1756-0500-1-54.
27. Snell C, Krypuy M, Wong EM et al. BRCA1 promoter methylation in peripheral blood DNA of mutation negative familial breast cancer patients with a BRCA1 tumour phenotype. Breast Cancer Res 2008; 10 (1): R12. doi: 10.1186/bcr1858.
28. Liu W, Gong J, Hu J et al. Quantitative assessment of AKAP12 promoter methylation in human prostate cancer using methylation-sensitive high-resolution melting: correlation with gleason score. Urology 2011; 77 (4): 1006.e1–1006.e7. doi: 10.1016/j.urology.2010.12. 010.
29. Dimitrakopoulos L, Vorkas PA, Georgoulias V et al. A closed-tube methylation-sensitive high resolution melting assay (MS-HRMA) for the semi-quantitative determination of CST6 promoter methylation in clinical samples. BMC Cancer 2012; 12 : 486–497. doi: 10.1186/1471-2407-12-486.
30. Mastoraki S, Chimonidou M, Dimitrakopoulos L et al. A rapid and accurate closed-tube methylation-sensitive high resolution melting analysis assay for the semi-quantitative determination of SOX17 promoter methylation in clinical samples. Clin Chim Acta 2015; 444 : 303–309. doi: 10.1016/j.cca.2015.02.035.
31. Yang X, Dai W, Kwong DL et al. Epigenetic markers for noninvasive early detection of nasopharyngeal carcinoma by methylation-sensitive high resolution melting. Int J Cancer 2015; 136 (4): E127–E135.
32. Liu W, Guan M, Su B et al. Rapid determination of AKAP12 promoter methylation levels in peripheral blood using methylation-sensitive high resolution melting (MS-HRM) analysis: application in colorectal cancer. Clin Chim Acta 2010; 411 (13–14): 940–946.
33. Xiao Z, Li B, Wang G et al. Validation of methylation-sensitive high-resolution melting (MS-HRM) for the detection of stool DNA methylation in colorectal neoplasms. Clin Chim Acta 2014; 431 : 154–163. doi: 10.1016/j.cca.2014.01.044.
34. Qiu C, Zhi Y, Shen Y et al. High-resolution melting analysis of HPV-16 L1 gene methylation: a promising method for prognosing cervical cancer. Clin Biochem 2015; 48 (13–14): 855–859. doi: 10.1016/j.clinbiochem.2015.05.006.
35. Jacquin E, Baraquin A, Ramanah R et al. Methylation of human papillomavirus type 16 CpG sites at E2-binding site 1 (E2BS1), E2BS2, and the Sp1-binding site in cervical cancer samples as determined by high-resolution melting analysis-PCR. J Clin Microbiol 2013; 51 (10): 3207–3215. doi: 10.1128/JCM.01106-13.
36. Candiloro IL, Mikeska T, Hokland P et al. Rapid analysis of heterogeneously methylated DNA using digital methylation-sensitive high resolution melting: application to the CDKN2B (p15) gene. Epigenetics Chromatin 2008; 1 (1): 7. doi: 10.1186/1756-8935-1-7.
37. Palecek E, Tkac J, Bartosik M et al. Electrochemistry of nonconjugated proteins and glycoproteins. Toward sensors for biomedicine and glycomics. Chem Rev 2015; 115 (5): 2045–2108. doi: 10.1021/cr500279h.
38. Palecek E, Bartosik M. Electrochemistry of nucleic acids. Chem Rev 2012; 112 (6): 3427–3481. doi: 10.1021/cr200303p.
39. Bartosik M, Palecek E, Vojtesek B. Elektrochemická analýza nukleových kyselin, bílkovin a polysacharidů v biomedicíně. Klin Onkol 2014; 27 (Suppl 1): S53–S60.
40. Campuzano S, Pedrero M, Pingarron JM. Electrochemical genosensors for the detection of cancer-related miRNAs. Anal Bioanal Chem 2014; 406 (1): 27–33. doi: 10.1007/s00216-013-7459-z.
41. Boon EM, Salas JE, Barton JK. An electrical probe of protein-DNA interactions on DNA-modified surfaces. Nat Biotechnol 2002; 20 (3): 282–286.
42. Bartosik M, Koubkova L, Karban J et al. Electrochemical analysis of a novel ferrocene derivative as a potential antitumor drug. Analyst 2015; 140 (17): 5864–5867. doi: 10.1039/c5an00958h.
43. Wang P, Han P, Dong L et al. Direct potential resolution and simultaneous detection of cytosine and 5-methylcytosine based on the construction of polypyrrole functionalized graphene nanowall interface. Electrochem Commun 2015; 61 : 36–39.
44. Deng H, Yang X, Yeo SP et al. Highly sensitive electrochemical methyltransferase activity assay. Anal Chem 2014; 86 (4): 2117–2123. doi: 10.1021/ac403716g.
45. Furst AL, Barton JK. DNA electrochemistry shows DNMT1 methyltransferase hyperactivity in colorectal tumors. Chem Biol 2015; 22 (7): 938–945. doi: 10.1016/j.chembiol.2015.05.019.
46. Furst AL, Muren NB, Hill MG et al. Label-free electrochemical detection of human methyltransferase from tumors. Proc Natl Acad Sci U S A 2014; 111 (42): 14985–14989. doi: 10.1073/pnas.1417351111.
47. Li W, Liu X, Hou T et al. Ultrasensitive homogeneous electrochemical strategy for DNA methyltransferase activity assay based on autonomous exonuclease III-assisted isothermal cycling signal amplification. Biosens Bioelectron 2015; 70 : 304–309. doi: 10.1016/j.bios.2015.03.060.
48. Muren NB, Barton JK. Electrochemical assay for the signal-on detection of human DNA methyltransferase activity. J Am Chem Soc 2013; 135 (44): 16632–16640.
49. Wu H, Liu S, Jiang J et al. A sensitive electrochemical biosensor for detection of DNA methyltransferase activity by combining DNA methylation-sensitive cleavage and terminal transferase-mediated extension. Chem Commun 2012; 48 (50): 6280–6282. doi: 10.1039/c2cc32397d.
50. Zhang L, Wei M, Gao C et al. Label-free electrochemical detection of methyltransferase activity and inhibitor screening based on endonuclease HpaII and the deposition of polyaniline. Biosens Bioelectron 2015; 73 : 188–194. doi: 10.1016/j.bios.2015.05.066.
51. Sina AA, Howell S, Carrascosa LG et al. eMethylsorb: electrochemical quantification of DNA methylation at CpG resolution using DNA-gold affinity interactions. Chem Commun (Camb) 2014; 50 (86): 13153–13156. doi: 10.1039/c4cc06732k.
52. Koo KM, Sina AA, Carrascosa LG et al. eMethylsorb: rapid quantification of DNA methylation in cancer cells on screen-printed gold electrodes. Analyst 2014; 139 (23): 6178–6184. doi: 10.1039/c4an01641f.
53. Wang P, Wu H, Dai Z et al. Picomolar level profiling of the methylation status of the p53 tumor suppressor gene by a label-free electrochemical biosensor. Chem Commun 2012; 48 (87): 10754–10756. doi: 10.1039/c2cc35 615e.
54. Bartosik M, Fojta M, Palecek E. Electrochemical detection of 5-methylcytosine in bisulfite-treated DNA. Electrochim Acta 2012; 78 : 75–81.
55. Epigenomics.com [homepage on the Internet]. Společnost Epigenomics [updated 2016 May 16; cited 2016 May 16]. Available from: www.epigenomics.com/en/ products-services/epi-procolon.html.
Labels
Paediatric clinical oncology Surgery Clinical oncologyArticle was published in
Clinical Oncology

2016 Issue Supplementum 4
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
- Metamizole at a Glance and in Practice – Effective Non-Opioid Analgesic for All Ages
- Metamizole vs. Tramadol in Postoperative Analgesia
- Spasmolytic Effect of Metamizole
- Metamizole in perioperative treatment in children under 14 years – results of a questionnaire survey from practice
Most read in this issue
- Úloha PD-1/PD-L1 signalizace v protinádorové imunitní odpovědi
- Nemalobuněčný karcinom plic – od imunobiologie k imunoterapii
- Nádorové buňky jako dynamický systém – molekulární a fenotypové změny v průběhu vzniku, progrese a šíření nádoru
- Nové metody studia metylace DNA – MS-HRM analýza a elektrochemie