
H-reflex – jeho role v neurofyziologii a klinice
H-reflex and Its Role in EMG Laboratory and Clinical Practice
H-reflex is the most extensively studied reflex in the electrophysiological literature. It is no longer considered to be strictly monosynaptic since it has been shown to contain a shorter monosynaptic and longer oligosynaptic component. It is widely used in EMG laboratories. The relative ease with which the H-reflex can be elicited makes it an attractive clinical tool. However, we must take into account the limitations to H-reflex examination in neurophysiology. This concerns the choice of appropriate methods used to elicit the H-reflex (correct location of stimulating and recording electrodes) and correct evaluation of amplitude and latency, which are influenced by many factors (including the phenomenon presynaptic of inhibition and post-activating depression). The H-reflex is not exclusively monosynaptic, it consists of monosynaptic and oligosynaptic pathways. This reflex is not an equivalent of tendon jerk reflex because it bypasses muscle spindle mechanisms. With correct interpretation, the H-reflex is a useful tool for diagnosing sensorimotor polyneuropathy, plexopathy, radiculopathy S1 and static nerve lesions. It has been used in sports medicine research to evaluate musculoskeletal injuries and can be used as a tool to assess the neurophysiologic mechanism underlying the recovery of walking after spinal cord injuries. H-reflex modulation is also used to monitor the degree of hypertonia in the study of spasticity.
Key words:
H-reflex – electromyography – presynaptic inhibition – post-activation depression – polyneuropathy – radiculopathy S1 – lumbar plexopathy
The author declares he has no potential conflicts of interest concerning drugs, products, or services used in the study.
The Editorial Board declares that the manuscript met the ICMJE “uniform requirements” for biomedical papers.
:
Z. Kadaňka Jr.
:
Neurologická klinika LF MU a FN Brno
:
Cesk Slov Neurol N 2017; 80(6): 641-646
:
Review Article
prolekare.web.journal.doi_sk:
https://doi.org/10.14735/amcsnn2017641
H-reflex je nejčastěji zmiňovaným reflexem v elektrofyziologické literatuře. Není již (jako dříve) považován za striktně monosynaptický reflex, protože bylo prokázáno, že se skládá z kratší monosynaptické a delší oligosynaptické složky. Je rutinně využíván v EMG laboratořích pro řadu předností (poměrně snadná a rychlá výbavnost), ale je třeba respektovat i všechny limitace, které vyšetřování tohoto reflexu provázejí. Jde zejména o správnou techniku vybavování reflexu (tj. správné umístění elektrod – stimulačních i snímacích) a též o kritické zhodnocení latence a amplitudy, které jsou ovlivněny mnoha faktory (zejména fenomény presynaptické inhibice a postaktivační deprese). Nejedná se o přesný elektrofyziologický korelát reflexu šlachy Achillovy, neboť při stimulaci neurologickým kladívkem jsou drážděny i jiné struktury než selektivně dostředivá Ia vlákna. Při správné interpretaci výsledků je H-reflex velmi užitečný nástroj v diagnostice zvláště senzomotorických polyneuropatií, proximálně lokalizovaných lézí charakteru plexopatie, radikulopatie S1 či postižení n. ischiadicus. V poslední době se hodnocení H-reflexu prosazuje i ve sportovní medicíně a při studiu lokomoce u pacientů s inkompletní míšní lézí. Modulace H-reflexu byva tež využivana k monitoraci stupně svaloveho hypertonu při studiu spasticity.
Klíčová slova:
H-reflex – elektromyografie – presynaptická inhibice – postaktivační deprese – polyneuropatie – radikulopatie S1 – lumbální plexopatie
Úvod
H-reflex je nejčastěji studovaný reflex v elektrofyziologické literatuře [1]. Je rutinně vyšetřován prakticky v každé elektromyografické (EMG) laboratoři, ale ne vždy si však uvědomujeme jeho význam a limitace, které vyšetření tohoto reflexu s sebou nese, vč. interpretace výsledků jeho měření. H-reflex může být vybaven až ze 20 svalů celého těla – horních i dolních končetin i obličeje, především ze svalů tonických. V běžné praxi se ovšem používá prakticky pouze H-reflex z m. soleus. Jeho klinické využití spočívá zejména v hodnocení proximálních segmentů periferního nervového systému (tj. plexů a nervových kořenů), které nejsou zahrnuty do standardního měření motorického a senzitivního vedení na končetinách, a je v těchto situacích dokonce citlivější než užití jiných komparativních metod (somatosenzorických evokovaných potenciálů [SEP], které vyšetří pouze senzitivní neurogram a mají rozdílnou aferentaci, či F vlny, jež nás informuje pouze o složce motorické) [2].
Historie a patofyziologie
V roce 1918 byl reflex poprvé popsán německým neurologem Johannem Hoffmannem. V roce 1951 ho Magladery charakterizoval jako striktně monosynaptický reflex, kdy se při submaximální elektrické stimulaci selektivně aktivují Ia aferentní vlákna svalových vřetének [3]. Signál se následně šíří do míchy, kde dochází k monosynaptické excitaci cílového motoneuronu a aktivaci příslušných svalových vláken jeho motorické jednotky. Tezi striktně monosynaptického charakteru H-reflexu však v roce 1984 zpochybnili Burke et al, kteří postulovali, že potenciál H-reflexu má příliš dlouhou vzestupnou fázi („rising phase“) na to, aby byl tvořen pouze monosynaptickou komponentou [4]. Následné studie na kočkách a poté i na lidech existenci oligosynaptických drah vláken typu Ia k homonymním svalům potvrdily [4,5]. V současné době je časná komponenta H-reflexu považována za monosynaptickou Ia dráhu, zatímco komponenta pozdní je dráha polysynaptická. Je totiž nepravděpodobné, že by elektrická stimulace smíšeného nervu vyvolala selektivně aktivaci pouze Ia vláken. Při stimulaci H-reflexu užíváme běžně takovou intenzitu submaximální stimulace, která vyvolá i malou M odpověď. Protože axony motoneuronů jsou v průměru menší než axony vláken skupiny Ia, je jasné, že při stimulaci periferního nervu mohou být aktivována i tenká vlákna Ib Golgiho aparátu a pravděpodobně i vlákna typu II ze svalových vřetének. Z toho je zřejmé, že všechny tyto komponenty se mohou spolupodílet na tvorbě H-reflexu [6]. H-reflex bývá často popisován jako elektrofyziologický korelát reflexu šlachy Achillovy, což není přesné, protože cesta šíření elektrického impulzu je v obou případech rozdílná (poklep kladívkem vyvolá excitaci i jiných struktur – především svalových vřetének – než selektivní submaximální stimulace elektrického impulzu, která svalová vřeténka obchází). Poměrně snadná výbavnost H-reflexu a jeho zdánlivě monosynaptický charakter bývají někdy v klinických studiích mylně interpretovány jako jediné měřítko excitability motorické jednotky. To však nemůže platit z následujících důvodů. Amplituda H-reflexu je sice ovlivněna množstvím neurotransmiteru, který je vyplaven z terminál Ia aferentních vláken, ale efekt uvolněného neurotransmiteru ovlivňují zejména dva fenomény: presynaptická inhibice a postaktivační deprese [7]. Mechanizmus presynaptické inhibice byl podrobně zkoumán již od počátku 60. let minulého století. Eccles et al prokázali u koček, že aktivace Ia vláken různých „vzdálených“ svalů vedla k presynaptické inhibici Ia aferentních vláken nervů zadní končetiny a k supresi H-reflexu [8]. U lidí byla presynaptická inhibice H-reflexu rovněž prokázána aktivací heteronymní svalové skupiny – např. stimulace n. peroneus communis vyvolala presynaptickou inhibici H-reflexu m. soleus [9]. Jiní autoři popsali, že i pasivní pohyb nohy vedl k poklesu ipsilaterálního H-reflexu m. soleus [10]. Je zajímavé, že suprese H-reflexu m. soleus byla popsána i u velmi mírné kontrakce tohoto svalu, což působí poněkud paradoxně, neboť právě volní aktivace soleu bývá využívána k facilitaci H-reflexu v EMG laboratořích (mechanizmus není zcela jasný, pravděpodobně je to způsobeno sníženou excitabilitou kortikospinálních neuronů danou zvýšenou intrakortikální inhibicí) [11]. Inhibiční mechanizmy H-reflexu však nejsou omezeny pouze na jednu končetinu. Cheng et al zjistili pokles jeho amplitudy i při pasivním pohybu druhé dolní končetiny [12]. Rytmické pohyby paží též vedou k fázické modulaci ipsilaterálního H-reflexu m. soleus [13]. Presynaptická inhibice pochází také z descendentních supraspinálních zdrojů [14]. Stimulace vestibulárních jader mozkového kmene vedla k aferentní depolarizaci Ia vláken na dolních končetinách [15]. Dokonce i pouhá mentální simulace pohybu (pacienti byli vyzváni, aby v duchu vyvíjeli tlak nohou k podložce) vedla k velkému poklesu excitability spinální reflexní dráhy, což ukazuje na to, že i kortikální struktury mohou vést k modulaci H-reflexu [16].
U postaktivační deprese se předpokládá, že je způsobena nedostatečným vyplavováním neurotransmiterů ze synaptických terminál, které byly nedávno aktivovány [17]. Tento jev přetrvává asi 8 s [18]. To je důležité si uvědomit, protože řada studií monitoruje H-reflex během pohybu, a tím je amplituda i výbavnost H-reflexu značně ovlivněna.
Technika vybavení H-reflexu z m. soleus
Pozice pacienta
Ideální poloha pro vybavení H-reflexu je sedící pacient s dolními končetinami flektovanými v kyčli a v kolenou a s fixovaným kotníkem ve 30˚ plantární flexi [19].Tím snížíme svalové napětí m. gastrocnemius oboustranně. Tento způsob vyšetření je však vhodný zejména k výzkumným studiím, v rutinní diagnostické praxi se spíše užívá poloha vleže na břiše (obr. 1).
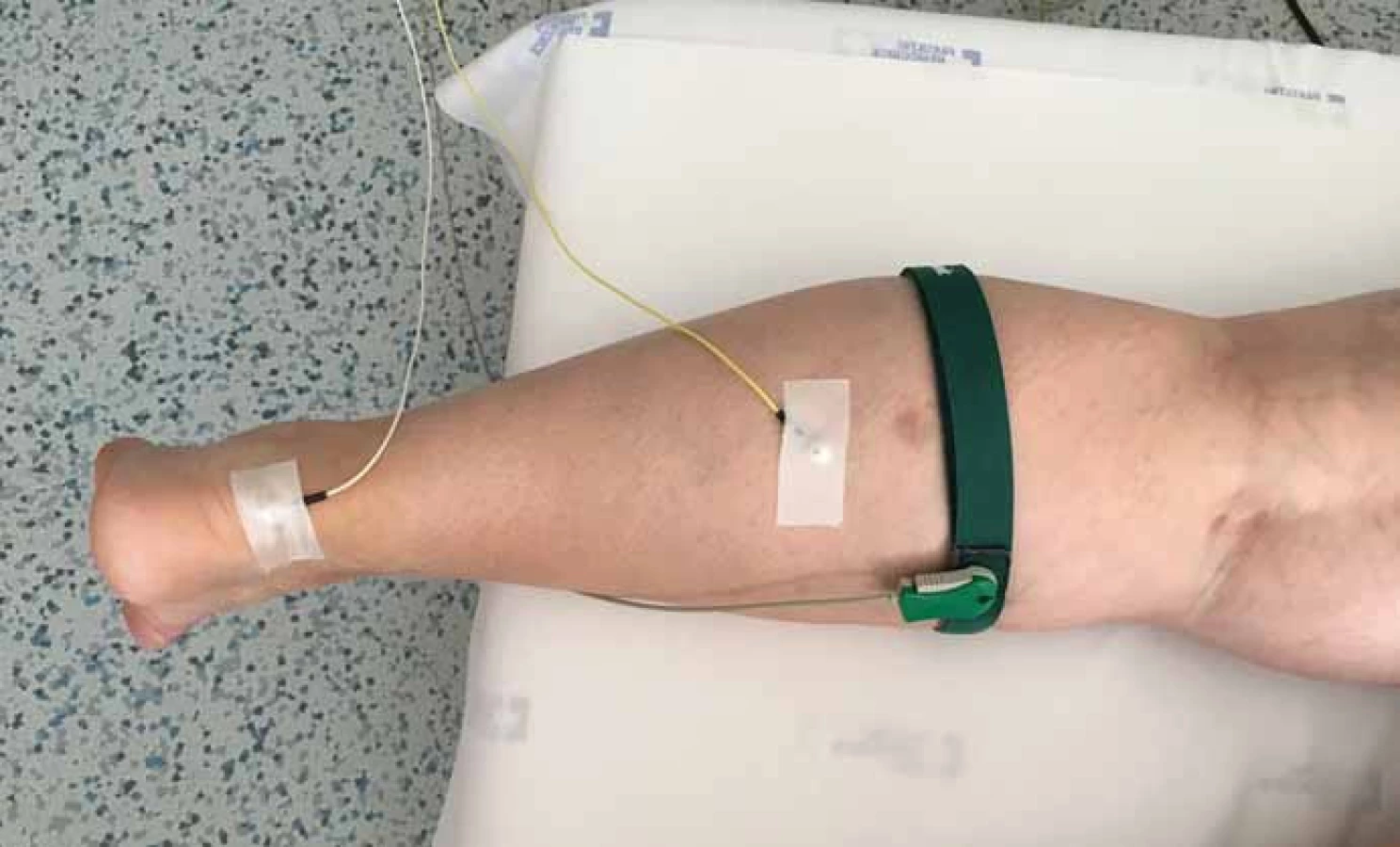
Stimulační elektrody
Existují dvě hlavní metody, jak stimulovat n. tibialis při vybavování H-reflexu. Při první z nich je katoda umístěna v popliteální jamce a anoda těsně nad patelou [19]. Druhou, častěji užívanou metodou, je stimulace s longitudinálně umístěnými elektrodami podél průběhu tibiálního nervu. Při využití této metody musíme dát pozor, zda není stimulován n. peroneus communis (stačí i vizuální kontrola, zda nedochází k dorziflexi nohy) [20]. První odchylka od izoelektrické linie musí být pozitivní M i H odpověď.
Šířka impulzu
O optimální délce trvání elektrického impulzu se diskutuje po řadu let. Dle současného konsenzu se považuje za optimální délka 1 ms, protože tím se maximalizuje rozdíl mezi vodivými vlastnostmi senzitivních a motorických nervů [6,21].
Frekvence stimulace
Pokud je pacient v klidu, tak se postaktivační depresi s jistotou vyhneme pouze tím, že použijeme nízkou frekvenci stimulace – jednou za 10 s. V běžné klinické praxi je však tato frekvence příliš nízká, a tak se jeví rozumnější kompromis se stimulací jednou za 3 – 5 s.
Umístění registračních elektrod
Hugon doporučoval, aby snímací elektrody měly velikost 1 cm2 a měly by být umístěny nad m. soleus na kůži – ve střední čáře zadní plochy lýtka asi 4 cm pod místo, kde se dvě hlavy m. gastrocnemius spojují s Achillovou šlachou, referenční elektroda je umístěna 3 cm distálně [19]. Zehr předpokládal, že by aktivní elektroda měla být umístěna na polovině vzdálenosti mezi popliteální jamkou a mediálním maleolem, referenční v oblasti Achillovy šlachy [7].
Měřené parametry
Latence
Latence se měří od začátku stimulu k první pozitivní odchylce křivky H-reflexu od základní linie, ne od začátku první negativity (obr. 2). To je podstatné, protože snímací elektroda na m. soleus není umístěna přímo nad motorickým bodem. Iniciální pozitivita H-reflexu je způsobena vedením vzruchu podél svalových vláken směrem k snímací elektrodě. Latence H-reflexu je závislá na výšce člověka a na jeho věku. Normální hodnoty jsou uvedeny v řadě normativních studií [22 – 24].
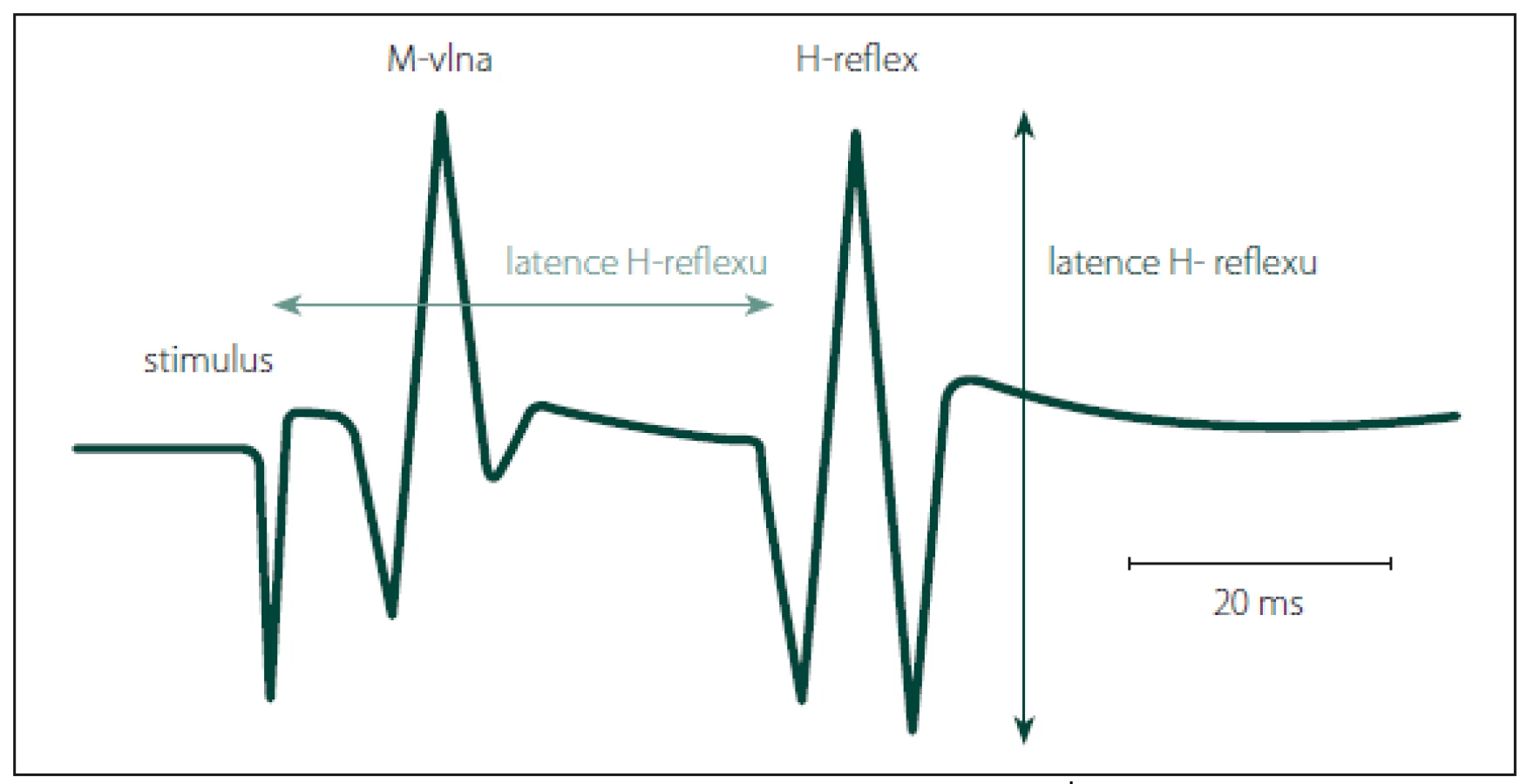
Amplituda
Amplituda H-reflexu je velmi variabilní a podléhá řadě vlivů (viz výše) (obr.3), je však užitečným měřítkem mezistranové diference při jednostranné patologii, především u radikulopatie S1.
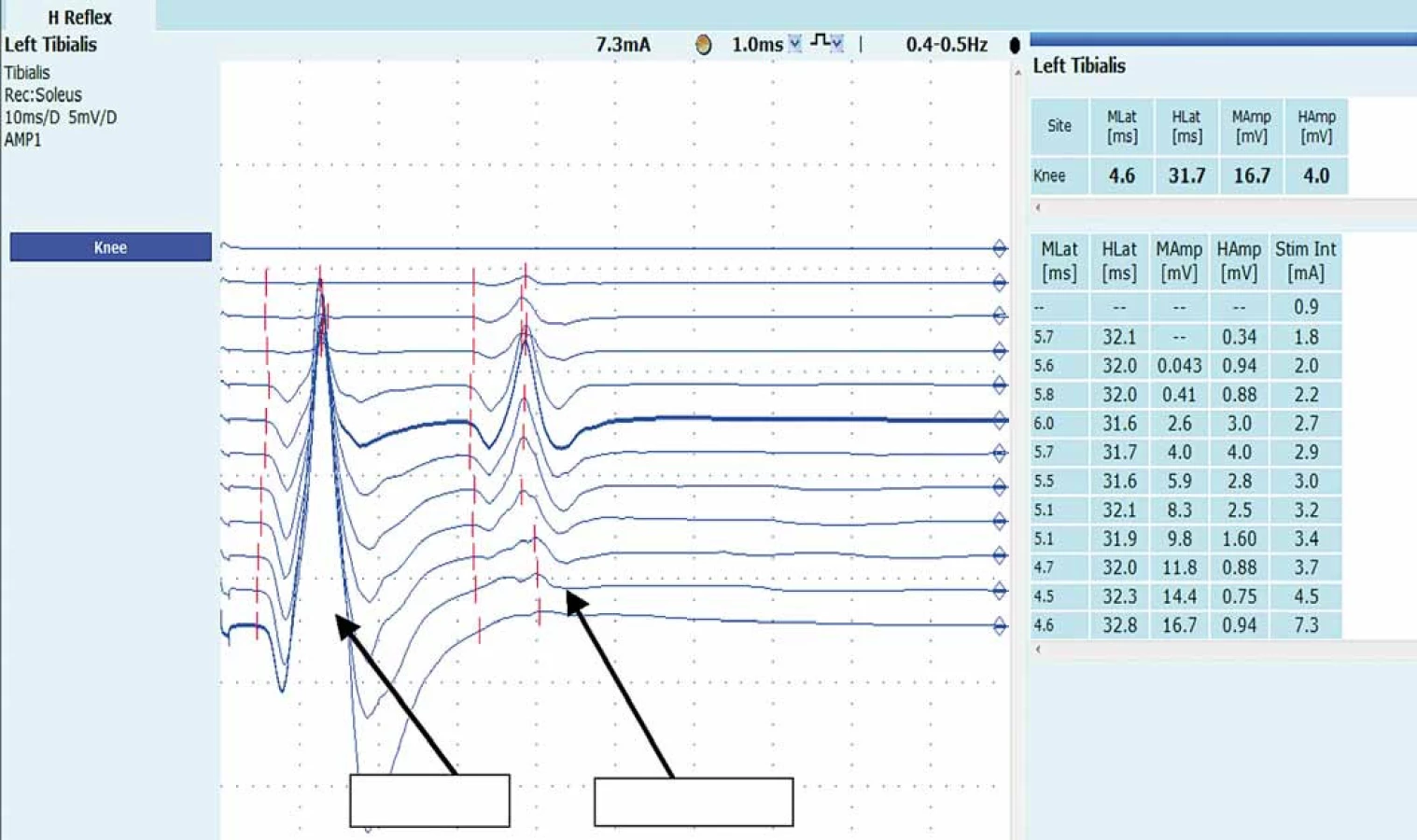
Nejvíce obhajovanou metodou normalizace amplitudy H-reflexu je její vyjádření jako procento Mmax. Nejprve změříme Mmax – ta je dána nejvyšší amplitudou M odpovědi, které dosáhneme při supramaximálním stimulu. Následně nastavíme intenzitu stimulace tak, aby dosahovala 10 % té hodnoty, kterou jsme vybavili nejvyšší amplitudu Mmax (např. pokud jsme Mmax dosáhli při intenzitě 30 mA, tak nyní budeme stimulovat 3 mA). Všechny následující H-reflexy budou tedy stimulovány touto konstantní intenzitou. Procento intenzity stimulace, které si vybereme, je arbitrárně stanoveno, většina autorů preferuje hodnotu mezi 10 a 25 % Mmax. Hmax je tak měřítkem maximální aktivace reflexu, což je dáno množstvím motoneuronů, které jsou schopny se aktivovat v dané situaci. Reprezentuje aktivaci celé motorické jednotky. Výhoda této metody spočívá v tom, že umožňuje hodnocení variabilní amplitudy H-reflexu prostřednictvím relativně stabilní amplitudy M odpovědi, která není ovlivněna míšními centry. Ale přes tyto předpoklady se ukázalo, že amplituda M odpovědi bývá ovlivněna různými okolnostmi při vyšetření (teplota, otok kotníku apod.) [25,26]. Předností této metody je relativní rychlost provedení.
Poměr Hmax/ Mmax (tzv. H/ M ratio) je další běžně užívaná metoda ke kvantifikaci amplitudy H-reflexu. Tj. porovnáváme nejvyšší amplitudu H-reflexu s nejvyšší amplitudou M odpovědi. Někdy bývá preferována před první (výše uvedenou) metodou, zejména v situacích, kdy může docházet k posunu stimulační nebo snímací elektrody. Nevýhodou metody je, že je méně citlivá k facilitaci či inhibici u vyšších amplitud H-reflexu. Byla nalezena korelace mezi H/ M ratio a stupněm spasticity. H/ M ratio je obrazem zvýšené excitability míšních alfa motoneuronů [27].
Význam H-reflexu v klinické praxi
U pacientů s polyneuropatií umožňuje vybavení H-reflexu porovnat vedení proximálními segmenty se segmenty distálními, má prediktivní hodnotu u diabetické polyneuropatie a poskytne nám více informací než běžné kondukční studie [28]. Čistě senzitivní abnormity přitom vytvářejí prolongaci latence jen mírnou, v řádu několika ms. Pokud je latence H-reflexu výrazně prodloužena, tak padá většinou na vrub postižení motorickému, protože výrazná aferentní abnormita by narušila přenos vzruchu natolik, že by k vybavení H-reflexu vůbec nedošlo [2].
Vybavení H-reflexu je též indikováno u pacientů s proximálně lokalizovanou lézí, zejména lumbosakrálních kořenů (u kořene S1 [29]), u pacientů s lumbální plexopatií (obr. 3) [30] či lézí n. ischiadicus. Je důležité mít normativní hodnoty vlastní laboratoře především pro mezistranové diference, např. Jankus et al uvádějí jako signifikantní rozdíl ≥ 0,4 ms [31]. Bilaterální prolongace latence H-reflexu m. soleus bývá považována za časný indikátor lumbální spinální stenózy či oboustranné radikulopatie S1 [32].

H-reflex je zvlášť užitečný v diagnostice akutní zánětlivé demyelinizační polyradikuloneuritidy, u které bývají nejčasněji postiženy proximální segmenty [33], jeho nevýbavnost bývá cenným podpůrným diagnostickým kritériem u Millerova-Fisherova syndromu (obr. 5) [34].
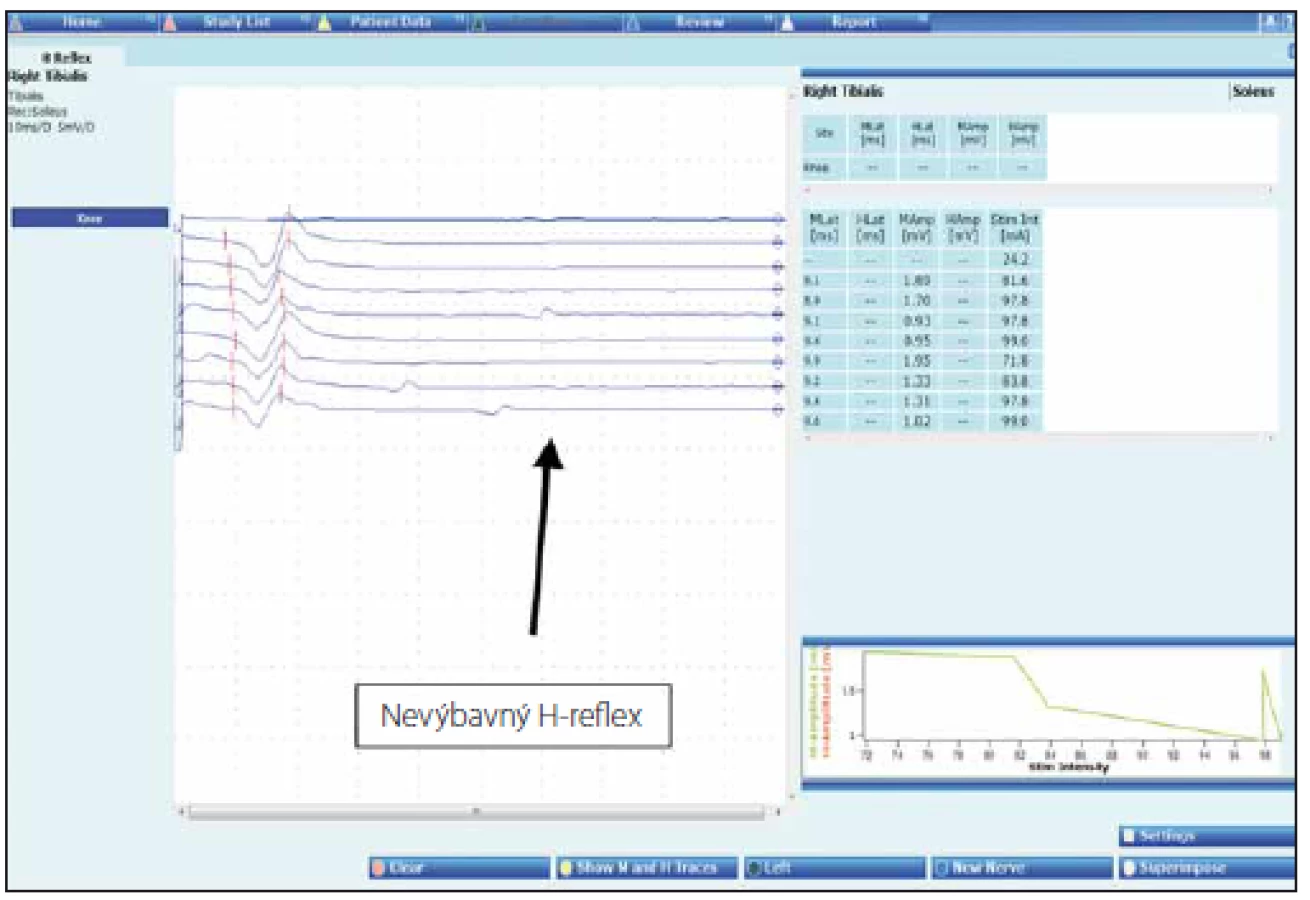
H-reflex bývá používán ve sportovní medicíně k hodnocení závažnosti muskoloskeletálních zranění, zhodnocení efektu jejich léčby (např. kryoterapií). Prokázalo se totiž, že při ochlazení končetiny dochází ke zvýšení amplitudy H-reflexu [35].
Modulace H-reflexu bývá též využívána k monitoraci stupně svalového hypertonu při studiu spasticity [36]. Například intratékálně podaný baklofen snižuje amplitudu H-reflexu již v prvních 15 min po podání [37]. Toho bývá využíváno i při hodnocení efektu intratékálně aplikovaného baklofenu – např. u dystonií, zejména u dětí [38].
Jednou ze slibných možností využití H-reflexu je při sledování adaptace míšních struktur. Již Chen a Wolpaw prokázali, že amplitudu H-reflexu lze u zvířat „natrénovat“ směrem nahoru i dolů procesem adaptace při dlouhodobém cvičení (tzv. up či down regulace H-reflexu) [39]. Následovala řada dalších studií u lidí, která potvrdila možnost adaptace H-reflexu i u nich. Např. signifikantně nižší amplituda H-reflexu byla zjištěna u členů Královského dánského baletu oproti trénovaným atletům apod. [40]. Tento fenomén (tj. up či down regulace) je v současné době v centru pozornosti řady klinických studií, kdy se jej autoři snaží využít ke zlepšení lokomoce u pacientů s míšní lézí. Například Chen et al uvádějí, že při up regulaci H-reflexu u myší (u kterých byla provedena inkompletní míšní léze) došlo k výrazné úpravě stoje a lokomoce [39]. Thompson et al prokázali, že u pacientů s inkompletní míšní lézí (po spinálním poranění), kteří podstoupili protokol „down regulace H-reflexu“, došlo k jasnému a výraznému zrychlení i úpravě lokomoce (což mělo vliv i na běžné denní činnosti) v porovnání s pacienty, kteří tento protokol neabsolvovali, a tedy ke snížení amplitudy H-reflexu nedošlo [41].
Závěr
H-reflex je velmi užitečný nástroj v EMG laboratoři, zejména v diagnostice polyneuropatií, proximálně lokalizovaných lézí charakteru plexopatie či radikulopatie. Musíme však respektovat všechna úskalí a limitace, které dané vyšetření provázejí. V poslední době se hodnocení H-reflexu prosazuje i ve sportovní medicíně, při studiu lokomoce u pacientů s inkompletní míšní lézí a k monitoraci stupně svalového hypertonu u spasticity.
Autor deklaruje, že v souvislosti s předmětem studie nemá žádné komerční zájmy.
Redakční rada potvrzuje, že rukopis práce splnil ICMJE kritéria pro publikace zasílané do biomedicínských časopisů.
MUDr. Zdeněk Kadaňka jr.
Neurologická klinika
LF MU a FN Brno
Jihlavská 20
625 00 Brno
e-mail: zdenek.kadanka@fnbrno.cz
Přijato k recenzi: 3. 7. 2017
Přijato do tisku: 16. 10. 2017
Sources
1. Misiaszek JE. The H-reflex as a tool in neurophysiology: its limitations and uses in understanding nervous system function. Muscle Nerve 2003;28(2):144 – 60. doi: 10.1002/ mus.10372.
2. Burke D, Gandevia SC, McKeon B. The afferent volleys responsible for spinal proprioceptive reflexes in man. J Physiol 1983;339(6):535 – 52.
3. Magladery JW, Porter WE, Park AM, et al. Electrophysiological studies of nerve and reflex activity in normal man. IV. The two-neurone reflex and identification of certain action potentials from spinal roots and cord. Bull Johns Hopkins Hosp 1951;88(6):499 – 519.
4. Burke D, Gandevia SC, McKeon B. Monosynaptic and oligosynaptic contributions to human ankle jerk and H-reflex. J Neurophysiol 1984;52(3):435 – 48.
5. Jankowska E, Johannisson T, Lipski J. Common interneurones in reflex pathways from group 1a and 1b afferents of ankle extensors in the cat. J Physiol 1981;310 : 381 – 402.
6. Lin CS, Chan JH, Pierrot-Deseilligny E, et al. Excitability of human muscle afferents studied using threshold tracking of the H reflex. J. Physiol 2002;545(2):661 – 9. doi: 10.1113/ jphysiol.2002.026526.
7. Zehr EP. Considerations for use of the Hoffmann reflex in exercise studies. Eur J Appl Physiol 2002;86(6):455 – 68. doi 10.1007/ s00421-002-0577-5.
8. Eccles JC, Schmidt RF, Willis WD. Presynaptic inhibition of the spinal monosynaptic reflex pathway. J Physiol 1962;161 : 282 – 97.
9. Iles JF. Evidence for cutaneous and corticospinal modulation of presynaptic inhibition of Ia afferents from the human lower limb. J Physiol 1996; 491(1):197 – 207.
10. McIlroy WE, Collins DF, Brooke JD. Movement features and H reflex modulation. II. Passive rotation, movement velocity and single leg movement. Brain Res 1992;582(1):85 – 93.
11. Misiaszek JE, Brooke JD, Lafferty KB, et al. Long-lasting inhibition of the human soleus H reflex pathway after passive movement. Brain Res 1995;677(1):69 – 81.
12. Cheng J, Brooke JD, Misiaszek JE, et al. Crossed inhibition of the soleus H reflex during passive pedalling movement. Brain Res 1998; 779(1 – 2):280 – 4.
13. Hiraoka K. Phase-dependent modulation of the soleus H-reflex during rhythmical arm swing in humans. Electromyogr Clin Neurophysiol 2001;41(1):43 – 7.
14. Chen XY, Wang Y, Chen Y, et al. Ablation of the inferior olive prevents H-reflex down-conditioning in rats. J Neurophysiol 2016;115(3): 1630 – 6. doi: 10.1152/ jn.01069.2015.
15. Rudomin P, Schmidt RF. Presynaptic inhibition in the vertebrate spinal cord revisited. Exp Brain Res 1999;129(1):1 – 37.
16. Bonnet M, Decety J, Jeannerod M, et al. Mental simulation of an action modulates the excitability of spinal reflex pathways in man. Brain Res Cogn Brain Res 1997;5(3):221 – 8.
17. Hultborn H, Illert M, Nielsen J, et al. On the mechanism of the post-activation depression of the H-reflex in human subjects. Exp Brain Res 1996;108(3):450 – 62.
18. Magladery JW, McDougal DB Jr. Electrophysiological studies of nerve and reflex activity in normal man. I. Identification of certain reflexes in the electromyogram and the conduction velocity of peripheral nerve fibres. Bull Johns Hopkins Hosp 1950;86(5):265 – 90.
19. Hugon M. Methodology of the Hoffmann reflex in man. In: Desmedt JE (ed). New Developments in Electromyography and Clinical Neurophysiology. Basel: Karger 1973 : 277 – 93.
20. Dumitru D, Amato AA, Zwartz MJ. Nerve conduction studies. In: Dumitru D, Amato AA, Zwarts M. (eds.) Electrodiagnostic Medicine. 2nd ed. Philadelphia: Hanley & Belfus 2001 : 159 – 223.
21. Panizza M, Nilsson J, Roth BJ, et al. The time constants of motor and sensory peripheral nerve fibers measured with the method of latent addition. Electroencephalogr Clin Neurophysiol 1994;93(2):147 – 54.
22. Lachman T, Shahani BT, Young RR. Late responses as aids to diagnosis in peripheral neuropathy. J Neurol Neurosurg Psychiatry 1980;43(2):156 – 62.
23. Sabbahi MA, Khalil M. Segmental H-reflex studies in upper and lower limbs of patients with radiculopathy. Arch Phys Med Rehabil 1990;71(3):223 – 7.
24. Buschbacher RM. Normal range for H-reflex recording from the calf muscles. Am J Phys Med Rehabil 1999;78(Suppl 6):S75 – 9.
25. Denys EH. M wave changes with temperature in amyotrophic lateral sclerosis and disorders of neuromuscular transmission. Muscle Nerve 1990;13(7):613 – 7.
26. Hicks A, Fenton J, Garner S, et al. M wave potentiation during and after muscle activity. J Appl Physiol 1989;66(6):2606 – 10.
27. Mayo M, DeForest BA, Castellanos M, et al. Characterization of involuntary contractions after spinal cord injury reveals associations between physiological and self-reported measures of spasticity. Front Integr Neurosci 2017;11 : 2. doi: 10.3389/ fnint.2017.00002.
28. Millán-Guerrero R, Trujillo-Hernández B, Isais-Millán S, et al. H-reflex and clinical examination in the diagnosis of diabetic polyneuropathy. J Int Med Res 2012;40(2):694 – 700. doi: 10.1177/ 147323001204000233.
29. Nishida T, Kompoliti A. Janssen I, et al. H reflex in S-1 radiculopathy: latency versus amplitude controversy revisited. Muscle Nerve 1996;19(7):915 – 7. doi: 10.1002/ (SICI)1097-4598(199607)19 : 7<915::AID-MUS19>3.0.CO;2-H.
30. Cho SC, Ferrante MA, Levin KH, et al. Utility of electrodiagnostic testing in evaluating patients with lumbosacral radiculopathy: An evidence-based review. Muscle Nerve 2010;42(2):276 – 82. doi: 10.1002/ mus.21759.
31. Jankus WR, Robinson LR, Little JW. Normal limits of side-to-side H-reflex amplitude variability. Arch Phys Med Rehabil 1994;75(1):3 – 7.
32. Kreiner DS, Shaffer WO, Baisden JL, et al. An evidence based clinical guideline for the diagnosis and treatment of degenerative lumbarspinal stenosis (update). Spine J 2013;13(7):734 – 43. doi: 10.1016/ j.spinee.2012.11.059.
33. Sudulagunta SR, Sodalagunta MB, Sepehrar M, et al. Guillain-Barré syndrome: clinical profile and management. Ger Med Sci 2015;21;13. doi: 10.3205/ 000220.
34. Dachy B, Deltenre P, Deconinck N, et al. The H reflex as a diagnostic tool for Miller Fisher syndrome in pediatric patients. J Clin Neurosci 2010;17(3):410 – 1. doi: 10.1016/ j.jocn.2009.06.014.
35. Kim KM, Hart JM, Saliba SA, et al. Relationships between self-reported ankle function and modulation of Hoffmann reflex in patients with chronic ankle instability. Phys Ther Sport 2016;17 : 63 – 8. doi: 10.1016/ j.ptsp.2015.05.003.
36. Karakoyun A, Boyraz İ, Gunduz R, et al. Electrophysiological and clinical evaluation of the effects of transcutaneous electrical nerve stimulation on the spasticity in the hemiplegic stroke patients. J Phys Ther Sci 2015;27(11):3407 – 11. doi: 10.1589/ jpts.27.3407.
37. Stetkarova I, Kofler M. Differential effect of baclofen on cortical and spinal inhibitory circuits. Clin Neurophysiol 2013;124(2): 339 – 45. doi: 10.1016/ j.clinph.2012.07.005.
38. Bonouvrié LA, Becher JG, Vles JS, el al. Intrathecal baclofen treatment in dystonic cerebral palsy: a randomized clinical trial: the IDYS trial. BMC Pediatr 2013;13 : 175. doi: 10.1186/ 1471-2431-13-175.
39. Chen XY, Wolpaw JR. Dorsal column but not lateral column transection prevents down-conditioning of H reflex in rats. J Neurophysiol 1997;78(3):1730 – 4.
40. Nielsen J, Crone C, Hultborn H. H-reflexes are smaller in dancers from The Royal Danish Ballet than in well-trained athletes. Eur J Appl Physiol Occup Physiol 1993;66(2):116 – 21.
41. Thompson AK, Wolpaw JR. Restoring walking after spinal cord injury: operant conditioning of spinal reflexes can help. Neuroscientist 2015;21(2):203 – 15. doi: 10.1177/ 1073858414
Labels
Paediatric neurology Neurosurgery NeurologyArticle was published in
Czech and Slovak Neurology and Neurosurgery

2017 Issue 6
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
Most read in this issue
- Brief Test of Verbal Memory Using the Sentence in Alzheimer Disease
- State-of-the-Art MRI Techniques for Multiple Sclerosis
- H-reflex and Its Role in EMG Laboratory and Clinical Practice
- When to Operate on Temporal Bone Fractures?