
Základy kognitivní, afektivní a sociální neurovědy
XI. Sociální rozhodování
Basics of cognitive, affective and social neuroscience.
XI. Social decision
The nature and means of decision has been of interest to philosophers since ancient Greece. A new scientific discipline has taken shape: neuroeconomics - which attempts to unify the advancements and research results of psychology, neuroscience and economy into a unified scope. Neuroeconomics has its supporters and vigorous opponents particularly among economists, some of who consider neuroeconomics dubious or irrelevant. Recently the first molecular genetic studies have begun to appear. The social decision described by the theory of expected utility and by the prospect theory has in its simplified point of view two tiers: the first phase is valuation, the second is a choice from a number of possibilities.
Key nodes of the “common neural currency” are
- the tegmentalis ventralis,
- pars compacta of the substantia nigra,
- nc. caudatus,
- the ventral striatum,
- orbitofrontal and medial prefrontal cortex.
Key nodes responsible for choice are
- parietal and
- lateral prefrontal cortex.
The evolutionary perspective says that human decision making is not guided by utility maximization but by operation of numerous decision rules shaped by selection to solve specific adaptive problems.
Key words:
social decision, neuroeconomics, expected utility theory, decision theory, evolutionary perspective.
Authors:
F. Koukolík
Authors‘ workplace:
Oddělení patologie a molekulární medicíny, Národní referenční laboratoř prionových chorob, Fakultní Thomayerova nemocnice s poliklinikou, Praha, Primář: MUDr. František Koukolík, DrSc.
Published in:
Prakt. Lék. 2011; 91(11): 632-638
Category:
Editorial
Overview
Povaha a způsoby rozhodování jsou předmětem zájmu filosofů od antického Řecka. Vyvíjí se nový obor: neuroekonomie. Ta se pokouší sloučit postupy a výzkumné výsledky psychologie, neurovědy a ekonomie do jednotného rámce. Má své zastánce i energické odpůrce, hlavně z řad ekonomů, někteří z nich považují neuroekonomii za pochybnou nebo irelevantní.
V současnosti se objevují první molekulárně genetické studie. Sociální rozhodování popisované teorií očekávaného užitku a teorií vyhlídek je zjednodušeně řečeno dvoustupňové: prvním stupněm je hodnocení, následuje volba mezi možnostmi.
Klíčovými uzly sítě „společné neuronální měny“ jsou
- area tegmentalis ventralis,
- p. compacta s. nigra,
- nc. caudatus,
- ventrální striatum,
- orbitofrontální, a
- mediální prefrontální kůra.
Klíčovými uzly sítě odpovídající za volbu jsou
- parietální, a
- laterální prefrontální kůra.
Evoluční perspektiva říká, že při sociálním rozhodování lidi nevede maximalizace subjektivního užitku, ale řada rozhodovacích pravidel selektovaných k řešení specifických adaptivních problémů.
Klíčová slova:
sociální rozhodování, neuroekonomie, teorie očekávaného užitku, teorie rozhodování, evoluční perspektiva.
Úvod
Povaha a způsoby rozhodování jsou předmětem zájmu filozofů od antického Řecka. Řešením otázky prosluli Blaise Pascal (1623–1662) a Daniel Bernoulli (1700–1782), Adam Smith (1723–1790), stejně jako utilitarističtí filozofové Jeremy Bentham (1748–1842) a John Stuart Mill (1806–1873), jenž byl i ekonomem a politikem.
Následoval rozsáhlý psychologický výzkum, od 60. let minulého století se přidala kognitivní neurověda, od 90. let neurověda afektivní. Sledovat činnost lidského mozku v průběhu rozhodování umožnilo zavedení funkčních zobrazovacích metod.
Formuje se neuroekonomie (28).
Ta se pokouší sloučit postupy a výzkumné výsledky psychologie, neurovědy a ekonomie do jednotného rámce. Má své zastánce i energické odpůrce, hlavně z řad ekonomů, někteří z nich považují neuroekonomii za pochybnou nebo irelevantní (přehled 3).
V současnosti se objevují první molekulárně genetické studie. Roe et al. (26) kupříkladu prokázal dva jednotlivé nukleotidové polymorfismy (SNIP, single nucleotide polymorphism) genu kódujícího alfa 4 nikotinový receptor (CHRNA4), které mají významný vztah k temperamentovému rozměru označenému harm avoidance, to je tendence vyhýbat se ohrožujícím vlivům (P1). Naproti tomu další temperamentový rozměr, novelty seeking, vyhledávání nového, má vztah k několika katechol-o-metytransferázovým SNIP. Postoj k ekonomickému riziku může mít vztah k několika SNIP vesikulárního monoaminového transportéru (VMAT2), signifikance této asociace je však slabá a její doložení nebo zamítnutí vyžaduje vyšetření větší kohorty.
Jinak řečeno: geny v nějakých mezích kódují stavbu a činnost funkčních systémů mozku a ta se v různých mezích projevuje nějakým druhem chování.
A naopak: cokoli se stane v sociokulturním poli, reflektuje vědomá a nevědomá činnost mozku včetně zpětnovazebného ovlivnění genové exprese.
Jsme součástí bio-psycho-sociálního (a historického) kontinua.
Některé vstupní pojmy
Blaise Pascal doporučoval vášnivým hráčům, aby odhadli pravděpodobnost různých možných vyústění hry. Rozhodování by mělo vycházet z výpočtu hodnoty v a pravděpodobnosti p, s níž tuto hodnotu lze získat.
Kvantitě (v*p) se říká očekávaná hodnota (expected value).
Dobrou předpovědí typu volby však není. Představte si, že jste v průběhu televizní hry vystaveni volbě mezi možností, že zcela jistě získáte 100 000 Kč a 50% možností zisku 500 000 Kč.
Většina lidí volí jistotu, byť je její očekávaná hodnota nižší. Podkladem tohoto rozhodnutí je nechuť riskovat (risk aversion). Jistých 100 000 Kč znamená pro chudého člověka mnoho, pro miliardáře je (ne vždy) částkou malou, je tedy teoreticky pravděpodobnější, že bude riskovat. Daniel Bernoulli proto usoudil, že rozhodnutí, neboli volba, záleží na subjektivní hodnotě u (utility, doslovně užitečnosti, užitku, přínosu) daného statku, respektive zboží. Volba je tedy ovlivněna subjektivně a záleží na očekávaném užitku (24; P2).
Předpovědi plynoucí z teorie očekávaného užitku však neplatí, jestliže je rozhodování zatíženo pocitem nejistoty, což odpovídá většině složitějších situací běžného života (1, 34). Rozhodování zatížené nejistotou je nutné odlišovat od rozhodování zatíženého rizikem. Při rozhodování zatíženém rizikem jedinec, jenž se rozhoduje, zná přesnou distribuci možných výsledků, například vsází-li na hod mincí nebo do loterie se známým počtem losů. Při rozhodování zatíženém nejistotou tuto informaci nezná, vyústění jen odhaduje (přehled 8).
Lidé špatně snášejí nejistotu, jestliže se rozhodují o možném finančním zisku, riskují však, to znamená, že lépe snášejí nejistotu, chtějí-li odvrátit možnou ztrátu. Chovají se tedy zdánlivě paradoxně. Tversky a Kahneman tyto zkušenosti vysvětlili teorií vyhlídek (prospect theory; 15, 32, 8), která předpokládá existenci funkčního systému umožňujícího vyhodnocování pravděpodobnosti a proměňujícího objektivní hodnotu v pocit subjektivního užitku. Teorie vyhlídek říká, že existuje proměnlivý referenční bod nebo souřadnice, k nimž vztahujeme pocit zisku nebo ztráty.
Neuronální podklady pocitu odměny
Desítky let experimentů se zvířaty ukázaly, že za možnost elektrické stimulace neuronálních oblastí mozku, které poskytují pocit slasti („centrální odměny“), vydrží zvířata elektrické šoky, snížení příjmu potravy a jsou s to se těžce fyzicky namáhat.
Jednou z klíčových oblastí je ventrální striatum, stejně klíčový je dopaminergní systém. Jestliže se ve ventrálním striatu blokuje vazba na dopaminergní receptor, jak prokázal starší experiment, hodnota stimulace coby odměny klesá (6). Velikosti odměny odpovídá kromě aktivity dopaminergních neuronů i činnost neuronů očnicové kůry, striata i zadní cingulární kůry (přehled 28).
Odměna aktivuje analogické neuronální sítě u lidí (obr. 1). Například Izuma et al. (12) nabídl zdravým dospělým lidem v průběhu dvou experimentů finanční odměnu, následně pak odměnu sociální. Finanční odměnu získávaly pokusné osoby v průběhu jednoduché hry.
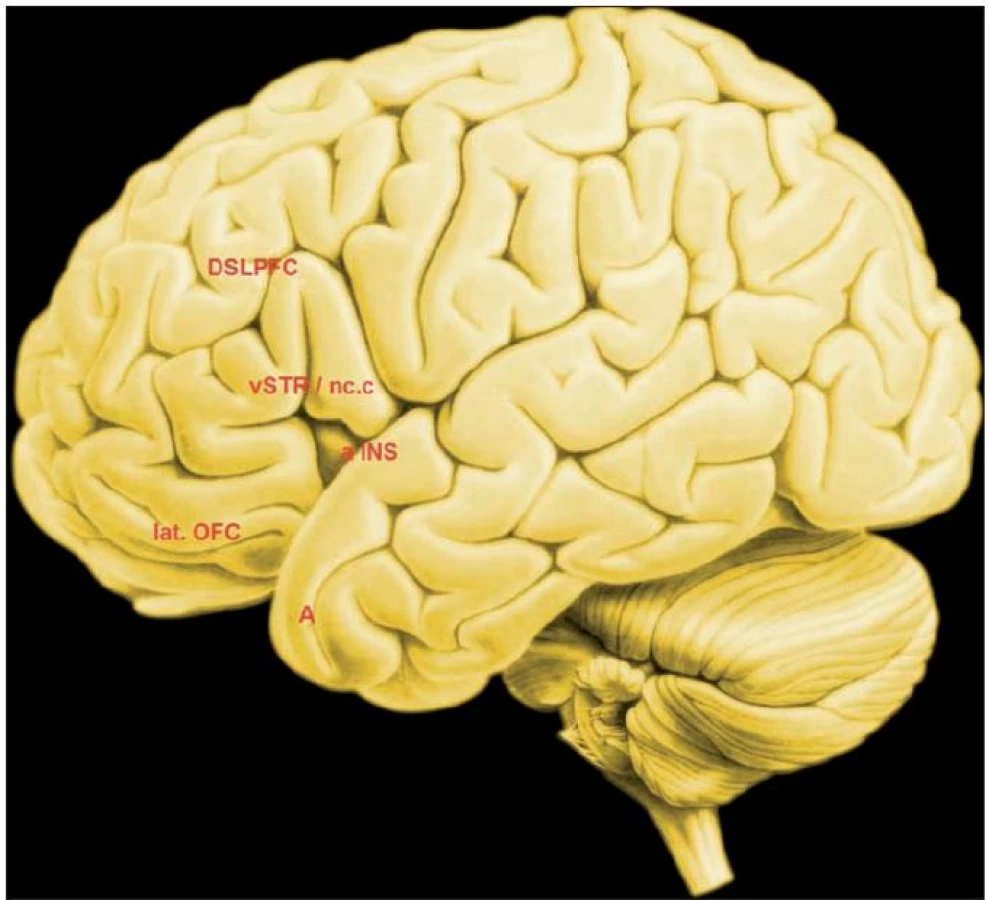
Složitější bylo zadání experimentu se sociální odměnou, ta byla dána ziskem dobré pověsti (reputace). Sociální odměna aktivovala neuronální síť „centrální odměny“, jejímž těžištěm je nc. caudatus a putamen, tedy striatum. Finanční odměna aktivovala síť, která se se s přechozí sítí rozsáhle překrývala, těžištěm aktivity bylo rovněž striatum.
Zink et al. (36) užila rovněž funkční magnetickou rezonanci v průběhu interaktivní hry. Pokusné osoby se při hře dostávaly do virtuální sociální interakce. Hráči přitom hráli se sociálně nadřazeným a podřazeným protihráčem. V jedné podobě hry byla sociální hierarchie stabilní, ve druhé byla nestabilní. Interakce se sociálně nadřazeným hráčem v porovnání s interakcí se sociální podřízeným hráčem při stabilní hierarchii aktivovala
- týlní a temenní kůru oboustranně,
- ventrální striatum,
- parahipokampální, a
- dorsolaterální prefrontální kůru.
Aktivace mozku hráčů se při nestabilní sociální hierarchii odlišovala. K oblastem aktivovaným hrou při stabilní hierarchii se přidala aktivace oblastí, které zpracovávají emoce, zejména amygdala, oblastí, které aktivuje sociální kognice, zejména mediální prefrontální kůra. Aktivace mozku při sociálním odměňování se opět podobala aktivaci při finanční odměně.
Hodnocení
Většina teorií vysvětlujících rozhodování dokládaných řadou experimentů uvedeného typu mluví o procesu, který je velmi zjednodušeně řečeno dvoustupňový.
Prvním stupněm je integrace nejrozlišnějších vlastností „nabídky“ do jednotné subjektivní hodnoty, těch může být v případě rozmanitých nabídek větší počet („mám si koupit zmrzlinu, lízátko, kousek dortu nebo obložený chlebíček?“ táže se subjekt), následuje volba.
Zajímavé je, že všechny rozmanité aspekty různých možností mozek převádí do jediné stupnice žádoucnosti, „společné neuronální měny“.
Klíčovými uzly „sítě společné neuronální měny“ jsou, jak plyne z experimentů se zvířaty i lidmi
- dopaminergní area tegmentalis ventralis,
- pars compacta černé substance,
- nc. caudatus,
- ventrální striatum,
- orbitofrontální, a
- mediální prefrontální kůra (obr. 2) (13).
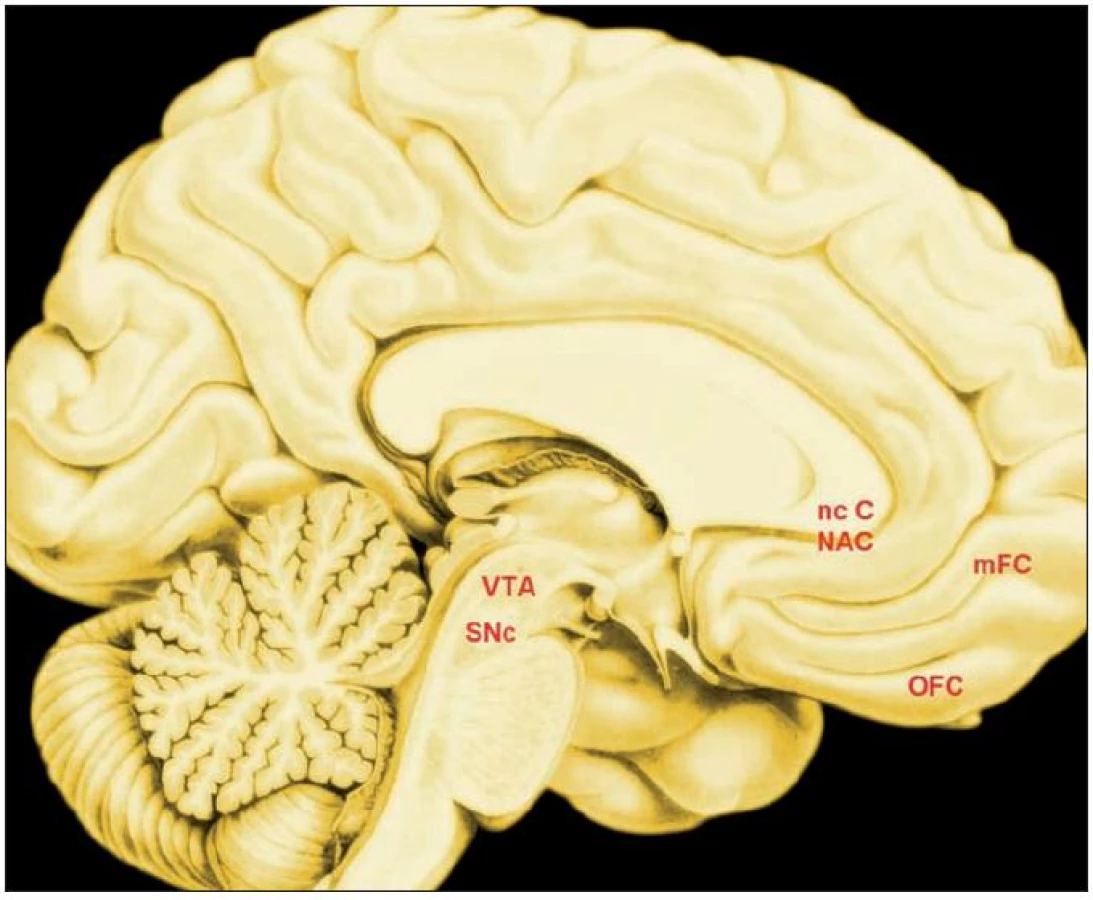
Jedním z dokladů je elegantní experiment s opicemi, jimž byla nabízena různá množství různých druhů ovocné šťávy, přitom se snímala aktivita neuronů area 13 orbitofrontální kůry. 80 % neuronů dané oblasti odpovídalo jedním ze tří způsobů:
- vyhodnocovaly nabídku, což bylo určeno porovnáním neuronální aktivity a typu chování pokusného zvířete,
- volily hodnotu, která byla nezávislá na druhu šťávy,
- volily chuť, která odpovídala druhu šťávy, tomu, který danému zvířeti chutnal.
Předpokládá se, že aktivita prvního typu odpovídá subjektivní hodnotě umožňující porovnávání a rozhodnutí mezi různými druhy nabídek (20).
V následné studii (21) autoři doložili, že je behaviorální odpověď zvířat konzistentní: jestliže zvíře dávalo přednost jablečné šťávě před hroznovou a této šťávě před čajem, dávalo přednost jablečné šťávě před čajem. Stejně konzistentní byla i odpověď vyšetřovaných neuronů.
Další experimenty ověřovaly činnost neuronů striata. U opic byly prokázány tři typy odpovědí, které aktivovaly stejné odpovědi neuronů orbitofrontálních. Činnost části těchto neuronů odpovídala hodnotě akce (action value neurons) bez ohledu na to, zda akci zvíře volilo, nebo nevolilo. Další skupina neuronů byla aktivní úměrně volené hodnotě a třetí byla aktivní v průběhu konkrétní akce (17).
Samejima et al (27) nechal pokusné opice rozhodnout, zda pohnout pákou vlevo nebo vpravo – podle směru pohybu dostávaly odměnu. Odměnou byl velký objem ovocné šťávy v porovnání s objemem malým. Elektrická aktivita třetiny modulovaných neuronů v putamen odpovídala hodnotě akce.
Experimenty s lidmi využívající funkční magnetickou rezonanci dospěly k podobným závěrům, byť je rozlišovací schopnost této metody podstatně nižší. Plassman et al. (23) nechal hladové osoby volit různé druhy potravy. Aktivita mediální prefrontální kůry odpovídala subjektivní hodnotě voleného jídla. Subjektivní hodnota volené potravy přitom odpovídá jak chuti, tak úvahám o významu dané potravy pro zdraví (9).
Kable a Glimcher (13) vyšetřovali aktivitu mozku v průběhu volby mezi okamžitou a odloženou finanční odměnou. Výše bezprostřední odměny byla stálá. Výše odložené odměny se měnila s časovou lhůtou. U lidí tak byla stanovena subjektivní diskontní funkce – čím delší byla doba, za kterou je možné odloženou odměnu získat, tím (obvykle) méně po ní lidé touží. Pokles subjektivní hodnoty odložené odměny odpovídal zejména aktivitě mediální prefrontální kůry a ventrálního striata.
Jinak řečeno: čím jsou lidé impulzivnější, a tím víc je zajímá bezprostřední odměna, tím strmější pokles aktivity obou zmíněných oblastí.
U trpělivějších lidí byl pokles méně výrazný. Neuronální aktivita obou oblastí tedy kóduje subjektivní hodnotu jak bezprostřední, tak odložené odměny v podobě společné „neuronální měny“, která bere v úvahu dobu, za kterou odměna nastane.
Lidské chování může charakterizovat jak nechuť vůči ztrátě, tak nechuť vůči nejednoznačnosti (ambiguity aversion). Kupříkladu Tom et al. (31) sledoval činnost lidského mozku v průběhu hry, v níž existovala možnost finančního zisku a ztráty v poměru 1 : 1. Aktivita ventromediální prefrontální kůry rostla úměrně zisku a klesala úměrně ztrátě. Poměr aktivity mozku v průběhu zisku k jeho aktivitě v průběhu ztráty odpovídal míře, v níž možnost ztráty ovlivňovala u daného jedince rozhodnutí víc než možnost zisku. Aktivita klesala rychleji úměrně rostoucí ztrátě u pokusných osob, jejichž vlastností byla nechuť ke ztrátě.
Levy et al. (18) vyšetřoval činnost mozku lidí, kteří se rozhodovali mezi pevně daným ziskem a hrou, která buď byla riskantní (takže pravděpodobnost různých vyústění byla známa), nebo byla víceznačná (takže pravděpodobnosti jednotlivých vyústění známy nebyly). Aktivita ventromediální prefrontální kůry a striata odpovídala subjektivní hodnotě rozhodnutí riskantních i rozhodnutí ve víceznačné situaci.
Subjektivní hodnota je u zvířat i lidí výsledkem učení, které je závislé na dopaminergní signalizaci ze středního mozku, zejména z neuronů area tegmentalis ventralis (Tsai; 19) (obr. 3).
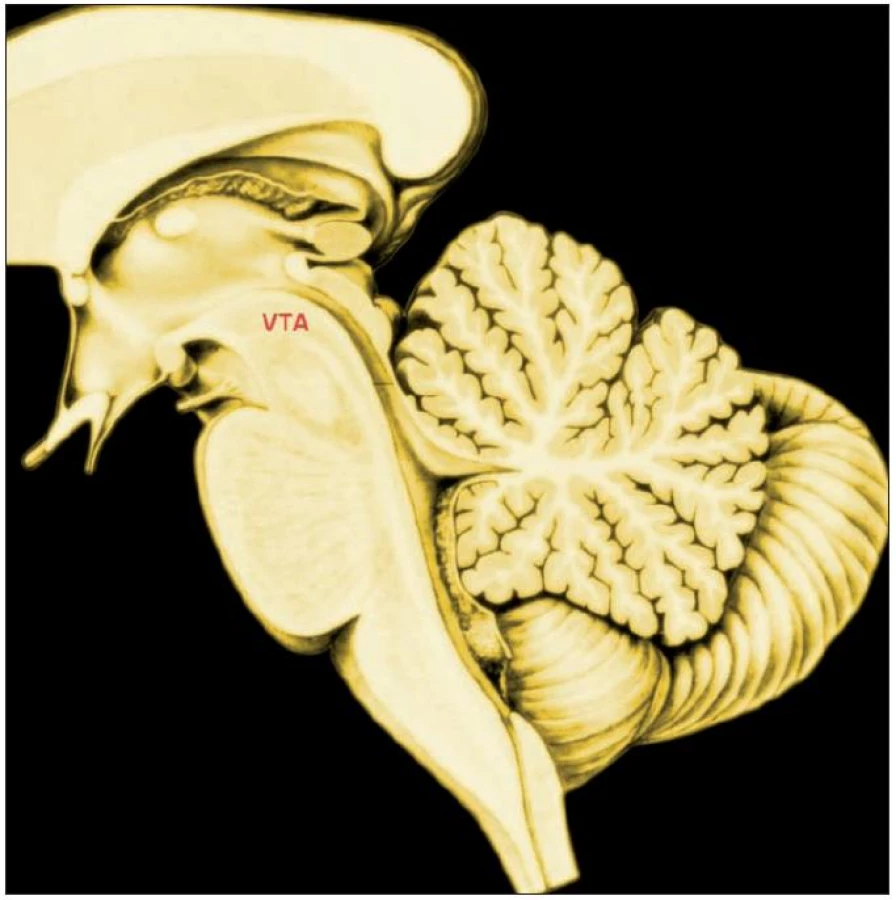
Subjektivní učení je podmíněno opakovaně aktualizovanou zkušeností. V průběhu experimentálního podmiňování dopaminergní neurony odpovídají na nečekanou odměnu, kromě toho odpovídají na první spolehlivou předpověď odměny po proběhlém podmiňování, na plně předpovězenou odměnu neodpovídají (proto je činnost dopaminergního systému jedním z podkladů temperamentového rozměru vyhledávání nového), jestliže se předpověď odměny nenaplní, jejich aktivita klesá (přehled 14, 29).
Zaghloul et al (35) zaznamenával u pacientů s Parkinsonovou chorobou, jimž byly implantovány elektrody v rámci stimulační terapie v průběhu učení elektrickou aktivitu neuronů s. nigra. Pacienti se učili, která ze dvou možností poskytuje vyšší pravděpodobnost hypotetické finanční odměny. Některé neurony s. nigra v porovnání s nečekaně záporným výsledkem rozhodnutí odpověděly zvýšenou aktivitou na nečekaně pozitivní výsledek rozhodnutí, zatímco se jejich aktivita při výsledku očekávaném neměnila.
Pessiglione et al. (22) doložil kauzální význam dopaminergní stimulace neuronů striata jak v průběhu učení, tak v průběhu předpovědi mylného signálu. Testované osoby dostávaly
- dopaminový prekurzor L-DOPA (L - 3, 4 dihydroxyphenylalanine),
- haloperidol (jenž je dopaminergním antagonistou) a
- placebo.
L-DOPA zlepšoval výběr odměny, možnost vyhnout se trestu neměnila.
Volba
Pouhé hodnocení k rozhodnutí nestačí. Nutná je volba některé možnosti. Její výběr z jejich množiny a převod do motorické akce jsou podmíněny činností sítě, jejímiž uzly jsou
- parietální, a
- zevní prefrontální kůra.
Plyne to ze sérií experimentů využívajících relativní dostupnost sítě odpovídající za iniciaci očních pohybů, například volních sakád. Rozhodnutí, že se opice a lidé podívají nějakým směrem, odpovídá aktivitě sítě tvořené laterální intraparietální oblastí (LIP), čelními poli očních pohybů (FEF, frontal eye fields, jsou součástí čelní kůry, leží v Brodmanově area 8, připočítává se oblast označovaná jako doplňkové pole očních pohybů, SEF, supplementary eye field) a horních hrbolků čtverohrbolí (přehled 14,15, 25).
Opět záleží na tom, zda jak jedinec, který se rozhoduje, zvládá riziko. Rostoucí riziko zvyšuje subjektivní hodnotu u lidí, kteří riziko vyhledávají (4), zatímco ji snižuje u lidí, kteří riziko nesnášejí. Tobler et al. (30) vystavil pokusné osoby dvěma druhům pokusné zátěže. V prvním experimentu volbu neměli, ve druhém volit mohli. Aktivita laterálních prefrontálních korových oblastí stoupala úměrně míře odměny za nepřítomnosti rizika v obou experimentech.
U lidí, kteří riziko vyhledávají, riziko aktivitu těchto oblastí zvyšovalo. U lidí, kteří mají k riziku nechuť, ji riziko snižovalo. Odpověď laterální prefrontální kůry tedy slučovala hodnotu volby a míru rizika. Aktivita striata naproti tomu odpovídala jen hodnotě, míra rizika ji neměnila.
Huettel et al. (11) dokázal, že rozhodnutí zatížené rizikem (nejistota se známými pravděpodobnostmi jednotlivých vyústění) nebo víceznačností (nejistota s neznámými pravděpodobnostmi jednotlivých vyústění) aktivují odlišné systémy. Zevní prefrontální kůru aktivovalo rozhodnutí zatížené víceznačností. Pokusné osoby, které dávaly přednost tomuto druhu rozhodování, charakterizovala nižší míra impulzivity. U lidí, kteří dávali přednost rozhodnutí zatíženému rizikem, se víc aktivovala zadní temenní kůra.
Tato dvojí disociace ukazuje, že rozhodování zatížené víceznačností není jen odlišnou podobou rozhodování zatíženého rizikem. Jde o dva druhy rozhodování podmíněné činností odlišných neuronálních sítí.
Podkladem nechuti ke ztrátě je aktivita amygdaly. Vzácná Urbachova-Wietheho choroba amygdaly poškozuje selektivně, aniž by poškozovala inteligenci. De Martino et al. (7) vystavil dvě pacientky experimentálním hrám o peníze, v nichž bylo možné vyhrávat a prohrávat. Schopnost obou pacientek odpovědět na proměny očekávaného zisku a míry rizika byla stejná jako u kontrolních jedinců. Ztráty však pacientky na rozdíl od kontrol nikterak neovlivňovaly.
K podobně iracionálnímu ekonomickému rozhodování vede poškození ventromediální prefrontální kůry (16).
Evoluční perspektiva
Ze zmíněné teorie Kahnemanovy a Tverského vyhlídek (prospect theory, 15,32) plyne, že by v průběhu rozhodování o zisku měli lidé dávat přednost možnostem spjatým s vyšším stupněm jistoty i v případě, že vyšší riziko znamená vyšší zisk. Čeština mluví o „hře na jistotu“, odborná terminologie užívá pojem efekt jistoty (certainty effect). A naopak: v průběhu uvažování o ztrátě by lidé měli dávat přednost riskantnější možnosti, byť by byla jistota spjata s menší ztrátou.
V běžném sociálním životě však někdy lidé tato pravidla nerespektují. Evoluční perspektiva říká, že lidi nevede maximalizace (subjektivního) užitku, ale řada rozhodovacích pravidel, která byla evolučně selektována k řešení specifických adaptivních problémů (2, 33). Hill a Buss (10) evoluční perspektivu dokládají experimentem, z něhož plyne, jak je lidské rozhodování spíše než maximalizací užitku ovlivňováno soutěží o postavení jedince na sociálním žebříčku.
Poznámky
Poznámka 1
SNIP – polymorfismus jednoho nukleotidu, je záměna jediného „písmene“ v genetickém kódu. Různí lidé tedy mohou mít dané sekvenci různé „písmeno“, což se v případě, že jde o polymorfismus v kódující sekvenci, nějakým způsobem projeví.
Sedmirozměrový model lidské osobnosti Cloningerovy skupiny (4, 5) nazývaný psychobiologický, protože je to jeden z prvních modelů, jenž uvedl do vztahu psychologické vlastnosti osobnosti s biologickými vlastnostmi mozku, nejprve popisuje čtyři rozměry osobnosti, které jsou různými druhy automatických odpovědí na podněty. Předpokládá, že druhy těchto automatických odpovědí jsou podmíněny z valné části dědičně.
Čtyři „rozměry“ osobnosti, jejichž podkladem jsou
- dědičně podmíněné,
- automatické,
- „podvědomé“, nebo také
-
„předpojmové“ odpovědi,
byly nazvány temperamentem.
Prvním rozměrem temperamentu je vyhledávání nového (novelty seeking). Dědičnost určí výběr podnětů, které dokáží jedince přimět ke zkoumání okolí, vyhledávání nových situací, impulzivnímu rozhodování nebo rychlým proměnám nálady.
Druhým rozměrem temperamentu je sklon vyhýbat se poškození (harm avoidance). Opět dědičnost vybere podněty, na jejichž základě jedinec některé druhy svého chování tlumí, začne se obávat budoucnosti nebo lidí, které nezná, nebo podněty, které rychle unavují.
Třetím rozměrem temperamentu je závislost na odměně (reward dependence). Dědičnost určí, jaké podněty nás přimějí, abychom v nějakém druhu chování pokračovali. Projevuje se například způsobem, jímž tvoříme sociální vazby nebo mírou závislosti na souhlasu druhých lidí s naším chováním.
Čtvrtým rozměrem temperamentu je vytrvalost (persistence). Je daná mírou vytrvalosti, s níž vzdorujeme únavě nebo tomu, že se činnost nedaří.
Rozsáhlá studie dvojčat ukázala, že dědičnost odpovídá za variabilitu čtyř temperamentových rozměrů v rozsahu mezi 50–65 %, přičemž tyto rozměry jsou vzájemně nezávislé. Rozdíly mezi nimi se projevují už u velmi malých dětí a do jisté míry předpovídají chování v průběhu dospívání i dospělosti.
Další tři rozměry lidské osobnosti nazval sedmirozměrový model charakterem. Jestliže byly temperamentové rozměry spíše dílem dědičnosti, pak jsou charakterové rozměry spíše výsledkem učení, tedy kulturních vlivů.
Lidé totiž proměňují vjemy a obsahy paměti na abstraktní symboly – pojmy. Lidská odpověď na podnět rozsáhle závisí na jeho pojmovém významu.
Naše automatické, „podvědomé“ nebo „předvědomé“ odpovědi spouštějící, udržující nebo tlumící nějaký druh chování jsou sice dány vrozenými temperamentovými vlastnostmi, nicméně je v každé chvíli výrazně ovlivňují pojmové významy podnětů.
Charakterové rozměry odpovídají tomu,
- do jaké míry se člověk cítí autonomním jedincem, tedy sám sebou,
- do jaké míry se cítí být součástí lidské společnosti, a
- do jaké míry se cítí být součástí celistvosti, vesmíru, celku, v němž je nějakým způsobem vše provázáno se vším.
Třemi rozměry charakteru jsou
- sebezaměření,
- ochota spolupracovat, a
- přesah.
Sebezaměření (self-directedness)
odpovídá schopnosti řídit a přizpůsobovat chování situačním proměnám tak, jak to odpovídá individuálně voleným cílům a hodnotám. Do značné míry odpovídá tomu, čemu se hovorově říká „síla vůle“. Sebezaměření se vyvíjí. Různé stupně sebezaměření jsou patrné již u dětí. Lidé s vysokým stupněm sebezaměření berou odpovědnost za svá rozhodnutí a činy na sebe, nepřesouvají je na lidi a okolnosti. Jejich život je naplňováním cílů, které mají tvořivý smysl. Mívají rozumnou míru sebedůvěry a umění řešit problémy. Dokážou brát sebe samé takové, jací jsou, mají rozumnou míru sebeúcty. Mají sklon lidem kolem sebe spíš důvěřovat, než vycházet z předpokladu, že je apriorní nedůvěra bezpečnější. Život je pro ně spíš naplňováním nějakého smyslu než uspokojováním bezprostředních potřeb a vyhýbání se střetům. Ve jménu dlouhodobých cílů dokáží uspokojení bezprostřední potřeby odložit.
Důležité je vědět, že:
nízká úroveň sebezaměření (neboli malá síla vůle) je společným jmenovatelem všech druhů poruch osobnosti.
Lidé s nízkou úrovní sebezaměření se často cítí jako oběť okolností, říkají, že jejich postoje převážně určují síly, které jsou mimo ně. Mají dojem, že jejich život nemá příliš velký smysl a žádný cíl, i pocit, že většina lidí je plodnějších, než jsou sami. Přejí si, aby byli hezčí a mocnější, než jsou lidé kolem nich. Jsou přesvědčeni, že mnohé z jejich vrozených vlastností jim znemožňují dosáhnout vyšší cíle, že sami „za nic nemohou.“
Ochota (nebo schopnost) spolupracovat (cooperativeness)
se objevuje u lidí společensky snášenlivých, vybavených značnou mírou soucitu i vciťování, respektujících práva i odlišnosti lidí jiných. Střety a problémy se snaží řešit způsobem, při němž obě strany něco získají. Domnívají se v souladu s židovsko-křestanskými a budhistickými mravními představami, že se zákony, které považují za přirozené, nedají porušit bez zlých následků pro jedince, společnost i přírodu. O těchto lidech se říká, že mají čisté srdce, na rozdíl od lidí vyhledávajících za každou cenu osobní výhody. Spolupracující lidé nebývají pomstychtiví.
Podobně jako v případě předchozího charakterového rozměru doprovází všechny druhy poruch osobnosti i nízká úroveň ochoty spolupracovat.
Sebe-přesah (self-transcendence)
je posledním charakterovým rozměrem. Sebe-přesah je chápán jako vědomí jednoty s celým bytím. Jedinec prožívá sám sebe jako část kosmického vývoje. Mluví se přijetí, pocitu sjednocení nebo duchovní jednoty s přírodou a jejím zdrojem. Schopnost sebe-přesahu se stupňovitě vyvíjí podobně jako předchozí charakterové rozměry. Objevuje se u vysokého podílu normálních lidí ze všech sociálních vrstev, nejen u světců a mystiků.
Lidé s vysokým sebe-přesahem často sdělují, že to, co právě dělají, je dokáže natolik pohltit, že se na chvíli cítí mimo čas a prostor. Dívají-li se na docela obyčejné věci, stane se, že mají pocit určitého zázraku, jakoby jev viděli poprvé, dětskýma očima. Prožívají chvíle radosti, v nichž mají náhlý pocit jednoty se vším, co je. Cítí se spjati se vším v přírodě, vše se jím zdá být jediným živým organismem. Někdy mluví o svém „šestém smyslu“, s nímž odhadují věci, které se teprve stanou. Mají dojem duchovní jednoty s jinými lidmi, kterou nedovedou vyjádřit slovy. Jindy mají pocit, že jejich život je veden duchovní silou větší, než je cokoli lidského.
Poznámka 2
Představte si, že se ráno rozhodujete, zda si na cestu do práce vezmete, nebo nevezmete deštník. Dejme tomu, že je pravděpodobnost deště 0,5, pravděpodobnost, že pršet nebude, je tedy stejná. V případě deště tedy sebou máte deštník, nebo ho nemáte. To samé v případě, že pršet nebude. Ve všech čtyřech případech přisoudíte možnému vyústění nějakou subjektivní hodnotu pohybující se od 0 do 100.
Nejhorší možnost 0 bodů, nejlepší 100 bodů.
Takže pro případ deště, při kterém máte deštník například 60 bodů, ve stejném případě a deštník nemáte 30 bodů.
A v případě, že pršet bude a nemáte deštník 0 bodů a že pršet nebude a deštník nemáte 100 bodů.
V tomto zadání je subjektivní očekávaná hodnota toho, že si deštník vezmete
[(0,5 x 60) + (0,5 x 30) ] = 0,45.
Stejná hodnota toho, že si deštník nevezmete, je
[(0,5 x 100) + (0,5 x 0) ] = 0,50 (Sanfey).
Vypadá to pedanticky, ale v mnoha životních situacích se toto jednoduché počítání vyplatí.
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř
prionových chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Sources
1. Bosaerts, P., Preuschoff, K., Hsu, M. The neurobiological foundations of valuation in human decision-making under uncertainty. In: Neuroeconomics. Decision making and the brain. Glimcher W., Camerer CF., Fehr E, et al. (vyd.). Academic Press 2009, p. 351-364.
2. Brandstätter, E., Gigerenzer, G., Hertwig, R. The priority heuristic: making choices without tradeoffs. Psychological Review 2006, 113, p. 409-432.
3. Clithero, J.A., Tankersley, D., Huettel, S.A. Foundations of neuroeconomics: from philosophy to practice. PLoS Biol. 6(11), e298. doi:10.1371/journal.pbio.0060298.
4. Cloninger, C.R., Svrakic, D.M., Przybeck, T.R. A psychobiological model of temperament and character. Arch. Gen. Psychiatry 1993, 50, p. 975-990.
5. Cloninger, C.R., Przybeck, T.R., Svrakic, D.M. et al. The temperament and character inventory: A guide to its development and use. Washington University Center for Psychobiology of Personality, St. Louis, Missouri, 1994.
6. Colle, L.M., Wise, R.A. Effects of nucleus accumbens amphetamine on lateral hypothalamic brain stimulation reward. Brain Res. 1988, 459, p. 361-368.
7. De Martino, B., Camerer, C.F., Adolphs, R. Amygdala damage eliminates monetary loss aversion. Proc. Natl. Acad. Sci. 2010, 107, p. 3788-3792
8. Fox, C.R., Poldrack, R.A. Prospect theory and the brain. In: Neuroeconomics. Decision making and the brain. Glimcher W., Camerer CF., ehr E, et al. (vyd.). Academic Press 2009, p. 145-173.
9. Hare, T.A., Camerer, C.F., Rangel, A. Self-control in decision-making involves modulation of the vmPFC valuation system. Science 2009, 324, p. 646-648.
10. Hill, S.E., Buss, D.M. Risk and relative social rank: positional concerns and risky shifts in probabilistic decision making. Evol. Hum. Behav. 2010, 31, p. 219-226.
11. Huettel, S.A., Stowe, C.J., Gordon, E.M. et al. Neural signatures of economic preferences for risk and ambiguity. Neuron. 2006, 49, p. 765-775.
12. Izuma, K., Saito, D.N., Sadato, N. Processing of social and monetary rewards in the human striatum. Neuron. 2008, 58, p. 284-294.
13. Kable, J.W., Glimcher, P.W. The neural correlates of subjective value during intertemporal choice. Nat. Neuroscience 2007, 10, p. 1625-1633.
14. Kable, J.W., Glimcher, P.W. The neurobiology of decision: consensus and controversy. Neuron. 2009,24, p. 733-745.
15. Kahneman, D., Tversky, A. Prospect theory: an analysis of decision under risk. Econometrica 1979, 47, p. 263-279.
16. Koenigs, M., Tranel, D. Irrational economic decision-making after ventromedial prefrontal damage: evidence from ultimatum game. J. Neuroscience 2007, 27, p. 951-956.
17. Lau, B., Glimcher, P.W. Value representation in the primate striatum during matching behavior. Neuron. 2008, 58, p. 451-463.
18. Levy, I., Rustichini, A., Glimcher, P.W. A single system represents subjective value under both risky and ambiguous decision-making in humans. Annual Society of Neuroscience Meeting, 2007, San Diego, CA.
19. Niv, Y., Montague, P.R. Theoretical and empirical studies of learning. In: Neuroeconomics: decision making and the brain. Glimcher PW, Camerer CF, Fehr E, et al., Academic Press, New York 2009, p. 331-351.
20. Padoa-Schioppa, C., Assad, J.A. Neurons in the orbitofrontal cortex encode economic value. Nature 2006, 441, p. 223-226.
21. Padoa-Schioppa, C., Assad, J.A. The representation of economic value in the orbitofrontal cortex is invariant for changes of menu. Nature Neuroscience 2008, 11, p. 98-102.
22. Pessiglione, M., Seymour, B., Flandin, G. et al. Dopamine-dependent prediction errors underpin reward-seeking behaviour in humans. Nature 2006, 442, p. 1042-1045.
23. Plassman, H., O´Doherty, J., Rangel, A. Orbitofrontal cortex encodes willingness to pay in everyday economic transactions. J. Neurosci. 2007, 27, p. 9984-9988.
24. Platt, M.L., Huettel, S.A. Risky business: the neuroeconomics of decision making under uncertainty. Nature Neuroscience 2008, 11, p. 398-403.
25. Rangel, A., Camerer, C., Montague, P.R. A framewok for studying the neurobiology of value - based decision making. Nature Reviews Neuroscience 2008, 9, p. 545-556.
26. Roe, B.E., Tilley, M.R., Gu, H.H. et al. Financial and psychological risk attitudes associated with two single nucleotide polymorphisms in the nicotine receptor (CHNA4) gene. PLoS ONE 4(8): e6704. doi:10.1371/journal.pone.0006704.
27. Samejima, K., Ueda, Y., Doya, K. et al. Representation of action-specific reward values in the striatum. Science 2005, 310, p. 1337-1340.
28. Sanfey, A.G., Lowenstein, G., Mc Clure, S.M. et al. Neuroeconomics: cross currents in research on decision making. Trends in Cognitive Neurosciences 2006,10, p. 108-116.
29. Tobler, P.N., Fiorillo, C.D., Schultz, W. Adaptive coding of reward value by dopamine neurons. Science 2005, 307, p. 1642-1645.
30. Tobler, P.N., Christoupoulos, G.I., O´Doherty, J.P. et al. Risk dependent reward value signal in human prefrontal cortex. Proc. Natl. Acad. Sci. 2009, 106, p. 7185-7190.
31. Tom, S.M., Fox, C.R., Trepel, C., et al. The neural basis of loss aversion in decision-making under risk. Science 2007, 315, p. 515-518.
32. Tversky, A., Kahneman, D. Advances in prospect theory: cumulative representation of uncertainty. J. Risk. Uncertain 1992, 5, p. 297-323.
33. Wang, X.T., Kruger, D.J., Wilke, A. Life history variables and risk taking propensity. Evolution and human behavior 2009, 30, p. 77-84.
34. Weber, E.U., Johnson, E.J. Decisions under uncertainty: psychological, economic, and neuroeconomic explanations of risk preference. In: Neuroeconomics. Decision making and the brain. Glimcher W., Camerer CF., Fehr E, et al. Academic Press 2009, p. 127-144.
35. Zaghloul, K.A., Blanco, J.A., Weidemann, C.T. et al. Human substantia nigra neurons encode unexpected financial rewards. Science 2009, 323, p. 1496-1499.
36. Zink, C., Tong, Y., Chen, Q. et al. Know your place: neural processing of social hierarchy in humans. Neuron. 2008, 58, p. 273-283.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2011 Issue 11
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
Most read in this issue
- Kalprotektin ve stolici
- Probiotika z pohledu praktického lékaře – klinické indikace pro podání probiotik, výsledky dotazníkového šetření mezi praktickými lékaři
- Diagnostika pľúcnej tromboembólie – úskalia CT angiografie
- Aktinická keratóza: poznatky o biologickom správaní a klinicko-patologických aspektoch ochorenia z pohľadu bioptika