
Základy kognitivní, afektivní a sociální neurovědy: XVII. Svobodná vůle
Basics of social cognitive and affective neuroscience: XVII. Free will
Voluntary action is perceived as essential to human nature. The neuroscience of free will refers to recent investigation of an ancient theological, philosophical, moral, psychological, neurological, psychiatric and legal question by neuroscientific tools. Does a rational agent exercise control over his actions, decisions and choices or is free will merely cognitive illusion? A large scale neuronal network exists for voluntary action. The primary motor cortex receives two classes of input. One from premotor – presupplementary and supplementary motor area which receives an input from basal ganglia and anterior prefrontal cortex and the second from early sensory cortices – parietal cortex – lateral premotor cortex. Frontopolar cortex forms and deliberates plans and intentions, presupplementary motor area in conjunction with other premotor areas prepares the action and generates the readiness potential the primary motor cortex becomes active. Free will is probably a biological trait. The philosophical notion of free will as existent or non-existent is probably a false dilemma. The more appropriate question about free will is not “if” we have a free will but “how much” free will do we have.
Key words:
free will, large scale neuronal network, biological trait.
Authors:
F. Koukolík
Authors‘ workplace:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
Published in:
Prakt. Lék. 2012; 92(4): 191-197
Category:
Editorial
Overview
Volní akci chápeme jako podstatnou složku lidství. Neurovědecké pojetí svobodné vůle odpovídá zkoumání prastaré teologické, filozofické, morální, psychologické, neurologické, psychiatrické a právní otázky neurovědeckými prostředky. Kontroluje racionální „agent“ své akce, rozhodnutí a volby, nebo je svobodná vůle pouhá kognitivní iluze? Podkladem volní akce je stavba a funkce neuronální sítě velkého rozsahu. Primární motorická kůra kůry dostává dva vstupy – jeden je z premotorické, presuplementární a doplňkové motorické kůry, která dostává informace z bazálních ganglií a přední prefrontální kůry, druhý z raných senzorických korových oblastí cestou temenní kůry a laterální premotorické kůry. Frontopolární kůra uvažuje o plánech a tvoří je, presuplementární kůra spolu s dalšími premotorickými oblastmi připravuje akci a je zdrojem přípravného potenciálu, pak se aktivuje primární motorická kůra. Svobodná vůle je pravděpodobně biologický znak a filozofické pojetí svobodné vůle jako buď existující, nebo neexistující může být falešné dilema. Správná otázka po svobodné vůli nezní, zda existuje, nebo neexistuje, ale kolik jí máme.
Klíčová slova:
svobodná vůle, neuronální síť velkého rozsahu, biologický znak.
Úvod
Požádám vás, abyste pohnuli, nebo nepohnuli ukazováčkem pravé ruky. Rozhodnutí je vaše. Ať jím pohnete, nebo nepohnete, máte pocit svobodné vůle: „Jsem to já, kdo se rozhodl ukazováčkem pravé ruky pohnout, nebo nepohnout“. Jde o fundamentální filozofický, teologický, morální, psychologický, právní a v poslední době i neurovědecký problém (příloha 1, 2, 3).
Je „agent“ (mlčenlivým předpokladem je v tomto případě zdravý dospělý člověk) svobodným „agentem“, záleží jen na jeho vůli, jak se rozhodne? Nebo je jeho svoboda pouhá kognitivní iluze (17, 23, 36, 21)? O pouhé intelektuální a filozofické rozjímání se touto otázkou nejedná: svobodná vůle je předpoklad odpovědnosti, právní i morální.
Onemocnění vůle
Pravděpodobně každý lékař v nějaké podobě, neurologové a psychiatři do podrobností, ví o řadě onemocnění, jimž se společně dá říci „onemocnění vůle“ – své stavy nejsou pacienti schopni volně zvládnout.
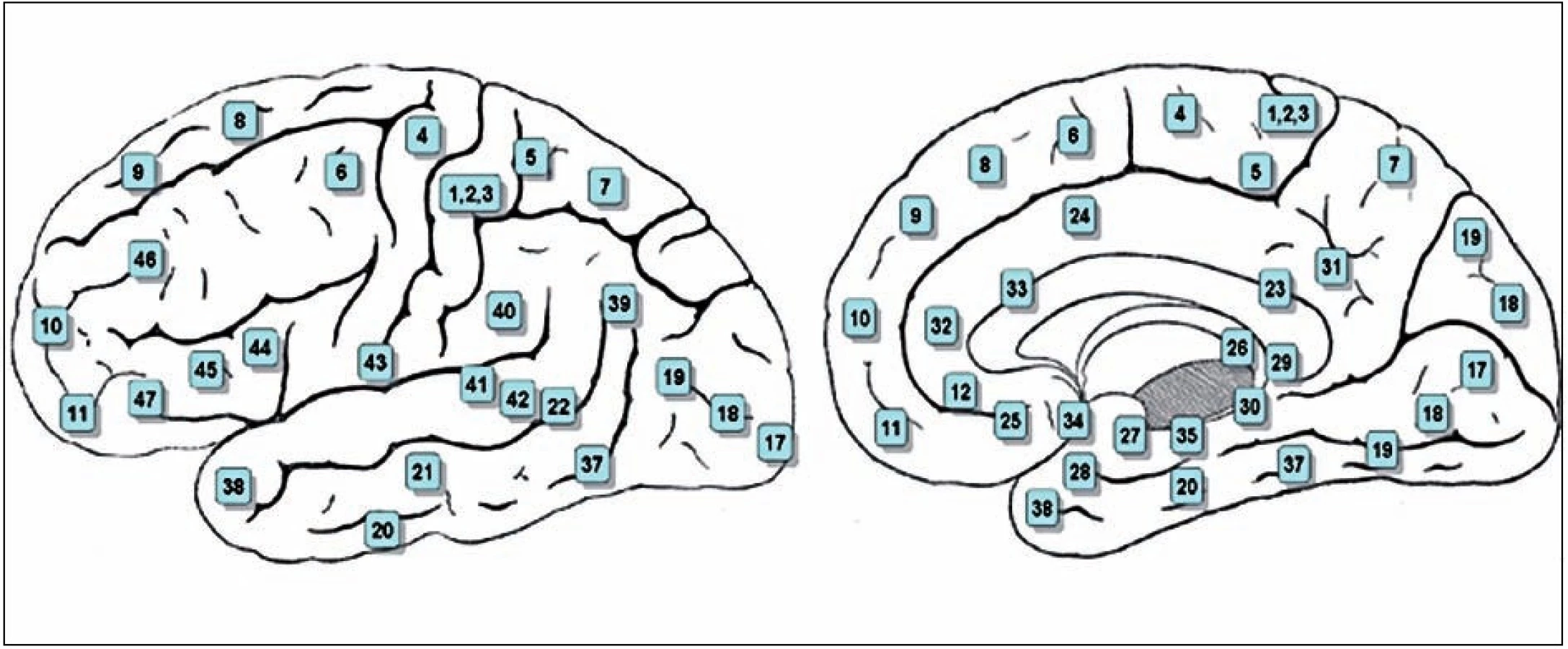
Příkladem je Huntingtonova nemoc. Pacienti mohou mít v počátečních fázích choroby pocit, že jejich mimovolní pohyby jsou volní, dále tiky, stejně jako Tourettův syndrom. Akinetický mutismus doprovází oboustranné poškození vnitřní čelní kůry v Brodmanově oblasti 6 (SMA, doplňková motorická kůra; obr. 2) a cingulární motorické kůry (část Brodmanovy area 24), zda je poškození těchto oblastí příčinou abulie, je nejasné. Každý lékař ví o Parkinsonově nemoci, mnozí vědí o syndromu odcizené, resp. anarchické ruky, která je důsledkem poškození doplňkové motorické kůry a premotorické kůry na vnitřní ploše hemisféry, kdy si postižená horní končetina „dělá, co chce“, sahá po okolí, rozepíná zapnuté knoflíky, pacienti na ní někdy sedí a na noc ji uvazují. Vysoký podíl populace postihuje v nějaké podobně obsedantně kompulzívní syndrom. ADHD (attention deficit hyperkinetic disorder) dříve zvaný lehká mozková dysfunkce, postihuje až 8 % školních dětí a v polovině případů přechází do dospělosti. Známé jsou disociativní stavy, například disociativní obrna, dále psychogenní tremor, syndrom fantomové končetiny, schizofreniky může týrat blud zevní kontroly, kdy mají nevývratný pocit, že jejich končetinami hýbe někdo cizí, nebo něco cizího (22).
Existuje tedy rozsáhlá skupina chorob, jejichž společným jmenovatelem jsou z hlediska této statě volně nekontrolovatelné pohyby. Funkčně anatomickým společným jmenovatelem je postižení neuronálních sítí velkého rozsahu, zejména motorického systému: prefrontální kůra, premotorická kůra – doplňková motorická kůra, dále obvodů prefrontální kůra – bazální ganglia – thalamus – další korové oblasti (20, 2, 35, 27). Součástí jsou projekce ze s. nigra a obousměrné propojení korových oblastí, thalamu a mozečku (22, 26).
Libetův experiment
V roce 1983 uveřejnil Benjamin Libet (1916–1927), fyziolog z kalifornské university, se svými spolupracovníky výsledky pokusu, který vešel do dějin fysiologie, neurovědy, práva i filozofie (31, 20) (obr. 1). Pokusné osoby seděly před hodinovým číselníkem. Na něm se pohybovala světelná skvrna, v jiné podobě pokusu hodinová ručička. Pokusné osoby se na ně dívaly a byly instruovány, aby se samy rozhodly, kdy pohnou rukou. Poté byly tázány, kde skvrna nebo ručička na číselníku byla, když tohle svobodné volní rozhodnutí udělaly. Tento okamžik byl označen písmene W (podle will =vůle). Tázány byly rovněž na okamžik, v němž se pohnuly, ten byl označen M (podle movement = pohyb).
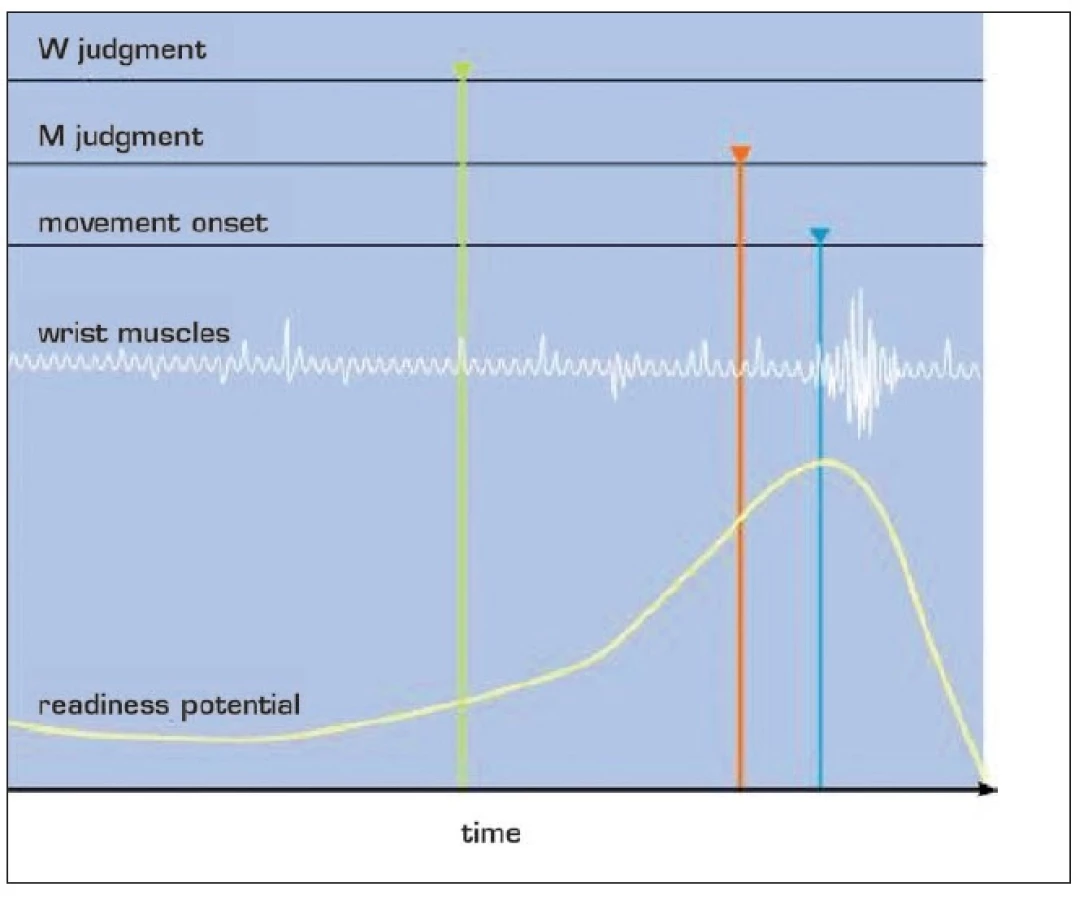
Začátek pohybu byl sledován elektromyograficky. Volní pohyby byly dvou typů: jeden byl promyšlený, druhý „spontánní, z rozmaru, jen tak“. Přesnost, se kterou lidé určují okamžik, v němž událost nastala, byla určována náhodnou elektrickou stimulací. Lidé oznámili její prožitek. Tento okamžik byl označen S, smyslový, senzorický podnět. V průběhu pokusu byla lidem snímána elektrická aktivita mozku, korové potenciály, které mají vztah k pohybu. To je vlna elektrické aktivity, která má několik složek.
První složkou je potenciál připravenosti, neboli Bereitschfatspotential (BP), v angličině readiness potentials, RP). Raná složka potenciálu připravenosti předchází vlastnímu pohybu o 1,5–0,4 s. Zdrojem je premotorická kůra a spolu s ní kůra doplňkové motorické oblasti, SMA, a to oboustranně. SMA je kůra „před“ primární motorickou kůrou, odpovídá area 6, zatímco primární motorická kůra, která je „za ní“, odpovídá area 4 Brodmanovy mapy korových polí lidského mozku (obr. 2). Potenciál připravenosti se objevuje i v temenní kůře.
Při promyšleně plánovaném pohybu se potenciál připravenosti objevil 1050 ms před pohybem. Při „pohybu z rozmaru“ přibližně 575 ms před pohybem. Okamžik W, tedy okamžik volního rozhodnutí, se objevil 200 ms před elektrickou aktivitou svalů odpovídajících pohybu. Vlastní pohyb si lidé uvědomili asi 90 ms před elektrickou aktivitou svalů. Jinak řečeno: Elektrická aktivita mozku předchází chvíli, v níž si uvědomíme své volní rozhodnutí, a to v případě plánovitého pohybu asi o 850 ms, v případě rozmarného rozhodnutí asi o 375 ms. Mozek tedy „ví“ o rozhodnutí dříve, než si to „uvědomí“.
Výsledky vyvolaly námitky a debatu, která svým způsobem pokračuje do těchto dní. Amerického filozofa Daniela Dennetta (nar. 1942) napadlo, že lidé přesouvají pozornost od svého záměru na hodiny, výsledkem je časový posun mezi pociťovaným rozhodnutím a vnímanou polohou ručičky na hodinách (11, 12). Prokázalo se, že přesuny pozornosti význam mají, nicméně výsledek pokusu odolal. Alfred R. Mele (nar. 1951), další americký filozof, si Libetův experiment vyzkoušel na sobě. Výsledku nedůvěřuje, má zato, že „vědomí záměru pohnout se“, tedy okamžik volního rozhodnutí, je nejistý, přinejmenším dvojznačný pocit, takže umístit jej v čase je problematické.
Libetův experiment byl však mnohokrát opakován, různými vědeckými skupinami, v různém uspořádání, výsledky odolaly. Činnosti mozku odpovídající za pohyb předchází okamžik, v němž máme pocit volního rozhodnutí.
Podobně jako další modifikace Libetova experimentu (19, 3) také experiment Soonovy skupiny (37), jenž užil funkční magnetickou rezonanci, prokázal, že závěry Libetovy skupiny platí: Lidé se dívali na obrázky písmen, které se objevovaly v rozestupu 500 ms. V některém okamžiku se spontánně rozhodli, zda stisknou tlačítko levým nebo pravým ukazováčkem. Tomu se v pokusu říkalo svobodná odpověď. Následně se podívali na obrazovku, která řekla: „Stiskem tlačítka určete okamžik, v němž jste se rozhodli ke svobodné odpovědi.“ Pokus dokázal, že aktivita prefrontální a temenní kůry předchází okamžiku, v němž vstoupí do vědomí až o 10 s (37).
Chvíli, v níž se rozhodujeme, nevnímáme, ale usuzujeme na ni
Podnětem pro pocit „teď jsem se rozhodl!“ by mohl být okamžik, v němž si lidé uvědomí, že odpověděli na podnět. Podnětem by tedy byl až vjem odpovědi na podnět, nikoli okamžik vlastního záměru „teď se rozhodnu“. Příčinnou souvislost mezi záměrem a akcí by to obracelo naruby. Banks a Isham (3) vytvořili iluzi, že k odpovědi na podnět došlo později, než k ní ve skutečnosti došlo. Testovaná hypotéza zněla: „Jestliže je okamžik, v němž vnímáme odpověď na podnět, rozhodující pro posouzení našeho záměru, pak zpoždění ve vnímaném času odpovědi posune i chvíli, kdy lidé mají za to, že se rozhodli.“
V prvním experimentu nabídli pokusným osobám klamnou zpětnou vazbu, zvukový signál, a to pípnutí, 5–60 ms po akci. Pípnutí oznamovalo, že pohyb začal později, než ve skutečnosti začal.
Podle předpokladu se rozhodnutí pokusných osob, tedy „svobodná volba“ začít akci, pohybovala v čase dopředu úměrně klamné zpětné vazbě. A to 5 ms po započetí svalové akce. Lidé se tedy již pohybovali a teprve potom měli dojem, že se k pohybu rozhodli. Ve druhém pokusu se lidé dívali na pohyby vlastní ruky, a to na videozáznamu – a ten buď byl, nebo naopak nebyl proti skutečnosti opožděný o 120 ms. Jestliže lidé sledovali falešnou signalizaci, opožděný videozáznam, udávali dobu vlastního rozhodnutí o 44 ms později, než tomu bylo v případě, kdy falešnou signalizaci nesledovali.
Lidé tedy na dobu, v níž se rozhodují, usuzují ze zdánlivého okamžiku, v němž se objevuje jejich akce (3–5, 28).
Situaci komplikuje skutečnost, že volní pohyby lze spustit podnětem, který subjektivně nevnímáme (9). Jestliže lidé vnímají sice dostačující, nicméně nepatrný podnět, zrakový nebo dotykový, rozliší ho. Jestliže je nepatrný podnět po velmi krátké době následován podnětem velkým, lidé podají zprávu jen o velkém podnětu, malý je „zpětně maskovaný“. Mozek zaregistruje a zpracuje podněty oba, ale do vědomí pronikne jen ten velký.
Experimenty s reakčním časem (38) doložily, že je stejný pro maskovaný podnět jako pro podnět nemaskovaný, snadno rozlišitelný. Lidé odpovídali na podněty, které subjektivně nevnímali, díky maskování jim totiž nevstoupily do vědomí. Za těchto okolností byl chod událostí: podnět, následovala odpověď na podnět, a teprve poté lidé vnímali, co se dělo. Sled událostí tedy nebyl podnět – vnímání – odpověď, což by byl „správný“ sled odpovídající svobodné vůli.
Pocit volního rozhodnutí závisí na pocitu příčinné souvislosti
Většina z nás má za to, že první je „myšlenka“, to znamená rozhodnutí, pak následuje pohyb. „Myšlenka“, akt svobodné vůle, je dle našeho pevného vědomí příčinou pohybu.
V experimentu ukazujícím, že pocit volního rozhodnutí k pohybu následuje až po elektrické aktivitě mozku, která je podkladem pohybu, lidé pohybovali prstem podle svého vlastního rozhodnutí. Do řady jejich pohybů náhodně zazněl zvukový signál ohlašující příkaz: „Stop!“. Pokusná osoba musela pohyb zastavit, jakmile uslyšela signál, byť si byla vědoma, že pohnout prstem chce. Vědomý záměr „pohnu prstem!“ se objevoval 1,42 s před započetím pohybu, ať už lidé prstem pohnuli, nebo pohyb na základě signálu zastavili.
Tato událost však následovala až poté, co se v jejich mozku objevil potenciál připravenosti. Čas, který uběhl do okamžiku, v němž si lidé uvědomili svůj záměr „pohnu prstem!“, byl příliš dlouhý na to, aby byl příčinou pohybu. Potenciál připravenosti se objevoval 2–8 s předtím. Vědomý pocit záměru „pohnu prstem!“ tedy není příčinou pohybu.
Jak pocit záměru, tak pohyb sám, jsou tedy paralelním, neboli souběžným výsledkem nevědomého zpracovávání informací (28, 20, 3, 4, 5, 19, 34).
Evolučně viděno je paralelita pocitu záměru a samotný pohyb pochopitelná. Okamžité pohyby na podnět mohou být nástrojem přežití. Buď přírodní výběr, nebo jiný základní mechanismus evoluce tedy „dal přednost“ tomuto systému.
Sebe-uvědomování, jáství, se v evoluci objevuje velmi pozdě, v plně rozvinuté podobě je zřejmě výsadou jen lidského druhu. Nadto se samo vyvíjí v průběhu života jedince. Práh vědomí – sebeuvědomování (neboli vědomí o vědomí jak svém, tak druhých lidí) překračují děti ve věku 36–48 měsíců. Sebeuvědomování vyrůstající z nevědomých procesů mozku se tedy s určitým zpožděním „dívá“ na to, jak probíhá motorická odpověď na zevní nebo niterný podnět. Tato odpověď rovněž vyrůstá z nevědomých procesů mozku (9, 25, 29, 30, 32, 33).
Volní akt jako prožitá zkušenost
Lidé pravděpodobně prožívají pocit vědomé svobodné vůle, jakmile si vlastní myšlení vyloží jako příčinu své akce. Filozof David Hume (24) první pochopil a poté v roce 1739, to mu bylo pouhých 28 let, napsal, že vůle není nic jiného, než „niterný dojem, který pociťujeme a jehož jsme si vědomi, jakmile vědomě dopřejeme vznik nového pohybu našeho těla nebo novému počitku naší mysli.“
David Hume pravděpodobně první na světě pochopil, že vůle není objekt spočívající v předmětech nebo lidech, ale spíše vjem plynoucí z trvalé souvislosti událostí. Zkušenost vůle jako něčeho osobního patrně plyne z priority, konzistence a výlučnosti myšlenek, které se akce týkají. Priorita říká, že myšlenka má ve správném časovém rozmezí předcházet akci. Konzistence mluví o tom, že myšlenka má akci odpovídat. Výlučnost sděluje, že myšlenka má být jediná zřejmá příčina dané akce (39).
Uspořádání většiny experimentů cílených na pochopení vztahu činnosti mozku a volní akce instruuje pokusné osoby třemi způsoby (20):
- pokusná osoba má vykonat nějakou akci, rozhoduje se, kdy ji vykoná;
- pokusná osoba má udělat nějakou akci v dané době, rozhoduje se, kterou akci z několika možností vybere;
- pokusná osoba se rozhoduje, zda akci provede, nebo neprovede.
Je pravda, že nejde o přirozené, ale o pokusné podmínky volního rozhodování, experimenty tohoto druhu rovněž nepostihují motivaci, důvod, proč volí pokusná osoba spíše jednu než jinou akci. Na druhé straně experimenty postihují to hlavní: Pokusné osoby musí samy vytvořit informace, které jsou nutné k provedení akce.
„Vykonavatelem“ a současně „posledním korovým článkem“ volní akce je primární motorická kůra (M1) v precentrálním závitu, levá řídí pohyby pravé poloviny těla, pravá pak pohyby levé poloviny těla. Informace předává prostřednictvím hybných neboli motorických drah příslušným oblastem páteřní míchy a ty je dodávají kosterním svalům. M1 je uzel ve dvou rozsáhlých sítích (obr. 3).
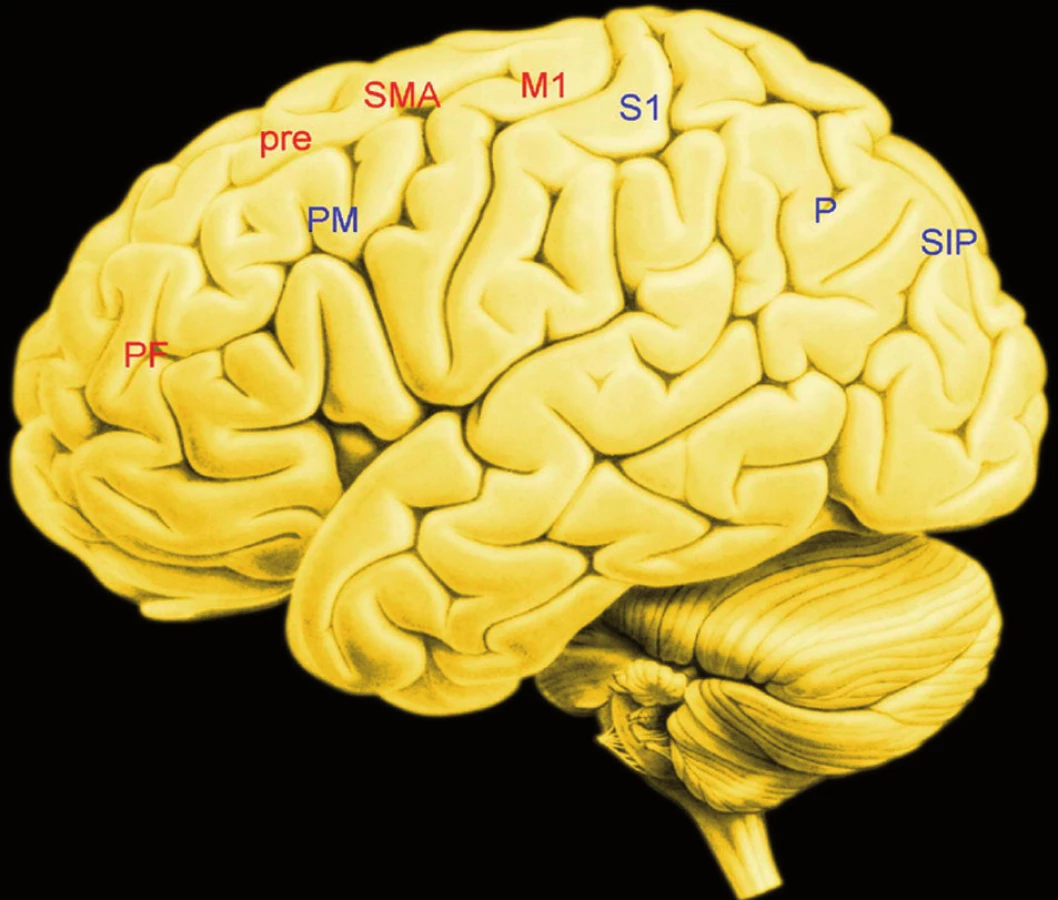
První, „přední“ neboli ventrální, je systém na obrázku 3 značený červeně. Jeho hlavními uzly jsou bazální ganglia nakupeniny nervových buněk v hloubi koncové části mozku, která informují prefrontální kůru, tedy nejpřednější část čelních laloků mozku, prefrontální kůra pak vysílá informace do kůry, která se jmenuje presuplementární a označuje se pre-SMA. Právě tato korová oblast se aktivuje výrazněji, jestliže se lidé rozhodují k pohybu samostatně, na základě vlastního volního rozhodnutí, než činí-li tak na základě instrukce, třeba tikání metronomu.
Pre-SMA sama je rovněž uzlem v další síti, jejímiž součástmi jsou kůra na pólu čelních laloků, kůra opaskového závitu a kůra, která je bezprostředně „před“ M1, takže se jí říká premotorická. Pre-SMA je zdrojem „potenciálu připravenosti“ (BP), v této oblasti se tedy rodí elektrická aktivita mozku směřující k akci, kterou si lidé uvědomí o něco později. Elektrická aktivita pre-SMA nevzniká „sama od sebe“ – něco ji musí spustit. Zřejmě ji spouští aktivita bazálních ganglií. Kromě jiného to plyne ze zkušenosti s pacienty stiženými Parkinsonovou nemocí. Elektrická aktivita předpovídající začátek pohybu se v bazálních gangliích objevuje 2 s před jeho skutečným začátkem, předchází tedy i elektrickou aktivitu odpovídající potenciálu připravenosti. Bazální ganglia sama jsou rovněž uzlem řady sítí včetně systému odměny – takže odtud mohou přicházet informace spouštějící volní akci.
Druhá síť, „zadní“, na obrázku 3 modře, jejíž informace se slučují v M1, je systém, přivádějící zprostředkované signály smyslových korových oblastí. Přivádí tedy například zrakové a současně prostorové informace. Jejími uzly jsou tedy smyslové korové oblasti – temenní laloky – zevní premotorická kůra a odtud projekce do M1. Volbě mezi různými cíli například odpovídá aktivita neuronů korové oblasti temenních laloků v oblasti sulcus intraparietalis.
Klíčová informace, která z řečeného plyne, zní: Neuronální systémy, jejichž činnost odpovídá volní akci, nejsou informační pyramida, na jejímž vrcholu by byla „vůle“. Jsou paralelně distribuovaným, neboli co do funkce souběžně uspořádaným systémem sítí, obvodů, „smyček“, jejichž činnost umožňuje pružné a inteligentní proměny chování odpovídající proměnám zevního světa (20).
V běžném jazyce je volní akce chápána jako niterná duševní volba. Ve skutečnosti je volní akce projev činnosti popsaných neuronálních obvodů, lze ji chápat jako rozhodování.
Přitom je nutné mít na mysli, že se rozhodování o vjemech liší od rozhodování o akcích. Rozhodnutí o vjemu je rozhodnutím o informacích, které vjem obsahuje. Rozhodnutí o akci vyžaduje tvorbu nových informací. To je proces, který je možné rozčlenit do čtyř fází (20):
- rané rozhodnutí „zda?“,
- rozhodnutí „co?“ – volba cíle,
- rozhodnutí „co?“ – volba pohybu,
- pozdní rozhodnutí „zda?“, konečná kontrola s předpovědí výsledku akce a případné veto.
První fázi předcházejí motivace, důvody pro akce. Akce ovlivní zevní prostředí, které zpětnou vazbou působí na motivace a důvody akcí.
Rané rozhodnutí „zda?“ Jestliže jsou jedincovy potřeby nasycené, lze jeho chování chápat jako soubor rutin, automatických programů, volně se rozhodovat nemusí. V opačném případě se jedinec rozhoduje, zda vůbec něco udělá. Rutinní zpracovávání informací nemusí dostačovat k rozhodnutí mezi dvěma nebo větším počtem možností při víceznačném podnětu. Může se objevit nový, mohutný „vegetativní“ podnět, například žízeň, hlad, strach nebo sexuální touha (tomu odpovídá aktivita kůry přední insuly) (5). Může se objevit libovolný podnět ženoucí jedince ke zkoumání prostředí.
Rozhodnutí „co?“ – volba cíle. Na volbě cíle se podílí zejména aktivita čelních laloků poté, co dostaly informace z bazálních ganglií, klíčovým uzlem je pre-suplementární kůra (pre-SMA).
Rozhodnutí „co?“ – volba příslušného pohybu se u lidí a pokusných opic testuje nejčastěji rozhodováním o prostorově rozmístěných cílech, například rozhodnutím o užití pravé nebo levé ruky, případně pohybu očí do levé nebo pravé strany.
Pozdní rozhodnutí „zda?“, konečná kontrola s předpovědí výsledku akce a případné veto: Informaci o provedení akce zpracovává několik neuronálních sítí, slučování jejich činnosti probíhá až před bezprostředním spuštěním. Mechanismy konečné kontroly umožňují nejen případnou úpravu akce na poslední chvíli, ale i její zastavení, vetování. U lidí se akce tohoto druhu často testují modelem Vpřed!/Stop! (Go/No-Go). Vyšetřovaní lidé očekávají nějaký signál, může jím být například křížek, který se objeví na obrazovce. Jakmile ho spatří, stisknou tlačítko, což se rychle naučí, protože podnět a odpověď se opakují. Jenže v malé části série se objeví signál říkající: „Tlačítko nestisknout!“ Vyšetřovaní lidé tedy musí utlumit odpověď, kterou se naučili. Zátěž testem Go/No-Go zvýší námahu přední části opaskového závitu v Brodmanově area 24. Jak klinické zkušenosti s lidmi postiženými neurologicky, tak experimenty s lidmi zdravými ukázaly, že se tato oblast podílí na sledování čili monitorování konfliktu. Konfliktem je zde náhlá změna situace: naučená odpověď „stisknout tlačítko jakmile se objeví signál!“ nutí stisknout, nový signál však říká „nestisknout!“.
Libetovy experimenty popisují druhou, výkonnostní polovinu dění. Nepopisují první, vstupní polovinu dění. Lidé, kteří přijdou coby pokusné osoby, vědí, co mají udělat, protože jim to experimentátoři řeknou. Jestliže jim to řeknou, pokusné osoby uloží tuto informaci do svých mozků, to znamená do „zadního obvodu“ svobodné vůle. Na pokusných osobách je, aby se rozhodly, kdy, učiní (nebo neučiní) jednoduchý hybný akt, například ohnou prst, nebo zápěstí. Tím užívají „přední obvod“ svobodné vůle.
Je pochopitelné, že mašinérie „nevědomí“, v experimentální situaci spuštěné povelem „rozhodněte se (svobodně)“, běží jako automat spuštěný tímto povelem podobně, jako se chystají hybné systémy sportovce na startovní výstřel. Výstřel pro sportovce je informace zvenčí, „rozhodl jsem se“ je startovní výstřel „zevnitř“, který vstoupí do běžícího automatického nevědomého dění – a může je zastavit. První polovinou událostí je tedy instruování pokusné osoby, která ví, co má udělat. Její „svobodné“ rozhodnutí spočívá v tom, zda to udělá, nebo neudělá, a jestliže to udělá, kdy to udělá. V obou případech implicitní systém komunikuje s automatickou, nevědomou senzoricko-motorickou mašinérií (1, 8, 25).
V biologii nemá smysl nic, není-li to ve světle evoluce
„V biologii nemá smysl nic, není-li to ve světle evoluce“, napsal 2 roky před smrtí Theodosius Dobzhansky (1900 až 1975) (10). Někteří soudobí biologové totiž mají za to, že svobodná vůle je „normální biologická vlastnost, nejde o dar ani tajemství“ (6, 7, 23, 32), což bude pravděpodobně překvapovat filozofy.
Modely, které se v této souvislosti studují, jsou únikové reakce. Příkladem předpověditelné odpovědi je C-startovní odpověď ryb. Ryby, které rozliší náhle zvýšený tlak z jedné strany těla, ohnou tělo do tvaru písmene C odvráceného od podnětu a snaží se bleskově uniknout. Vodnářka tykadlová (Erpeton tentaculatus), vodní had, vyvolává tuto předpověditelnou odpověď rybek pohybem zadní části těla, takže rybky obratem vyrazí přímo do její tlamy.
Předpověditelnou odpověď much dobře maskovaných v povrchovém krytu místa, kde pobíhají, podobně provokuje i lednáček bělokřídlý (Myioborus pictus). Mouchy vyletí, pták je snadno rozliší proti nebi, takže se stanou jeho kořistí.
Deterministické, předpověditelné chování však nemůže být evolučně stabilní. Živočich, který se chová předpověditelným způsobem, má nižší naději na přežití, než živočich, který se chová nepředpověditelně. Selekční tlak dává přednost nepředpověditelnosti. Variabilita chování je tudíž adaptivní vlastnost.
Jestliže ve dvou uvedených příkladech je chování živočichů předpověditelné, pak je příkladem horší předpověditelnosti chování švábů. V zadní spodní části těla mají cerci, dva vláskovité útvary (na každé straně jeden), což jsou čidla detekující pohyby vzduchu. Jakmile je čidla rozliší, spustí únikovou reakci, která šváby vede od místa, kde pohyb rozlišili. Úhel úniku ve vztahu ke směru vzdušného pohybu je však vysoce proměnlivý, tím je směr úniku pro predátora určitelný obtížně (13).
Filozofický spor o svobodnou vůli bývá dichotomický – strany sporu jsou buď zastánci determinismu, nebo indeterminismu. Pravděpodobně jde o falešnou dichotomii, neboli černobílý klam. Pozorování i experimenty ovlivňující chování zvířat ukazují, že jejich chování není zcela náhodné a přitom je špatně předvídatelné. Mozky zvířat a lidí lze chápat jako nestabilní nelineární systémy vysoce citlivé na nepatrné odchylky vstupních podmínek. Následné zesilování odchylek neboli amplifikace je příčinou případně exponenciálně odlišného výsledku činnosti stejných systémů, jejichž vstupní podmínky byly na počátku téměř stejné.
Experimenty s octomilkami ukazují, že jejich chování je daleko proměnlivější, než by odpovídalo proměnám činnosti neuronů, které zprostředkovávají reakci podnět – odpověď. Byť se v prostředí neděje nic, co by je nutilo měnit chování, chování proměňují. Každá z proměn je spontánní akce, z nitra systému, není odpovědí na spouštějící zevní podnět.
Jiným příkladem může být izolovaný nervový systém pijavky, který na základě neměnného elektrického podnětu někdy volí motorický program odpovídající plavání, jindy lezení. V tomto případě signály, které by mohly uniknout experimentátorově pozornosti, neexistují, proměny chování vznikají v systému samotném (18, 7, 23).
Chápat zdánlivě jednoduché živočichy jako biologické roboty by bylo zcela chybné. K pochopení složitosti jejich života nutná žádná mystika. Octomilky, stejně jako jiní živočichové a lidé, neznají řešení všech problémů předem. Nízká předpověditelnost jejich chování zvyšuje pravděpodobnost úniku před predátory a úspěchu v soutěži s příslušníky vlastního druhu; kromě toho hledají různá řešení nepředvídaných a nepředvídatelných problémů. Čím víc je „pasivní“ odpověď nahrazována akcí, tím větší pravděpodobnost evolučního úspěchu: přežití a potomstva.
Jestliže chápeme proměnlivost a malou předpověditelnost chování jako adaptivní znak, pak je nutné odlišovat odpovědi od akcí. Akce jsou druhy chování, u nichž nelze zjistit vyvolávací podnět, nebo je latence a mohutnost odpovědi natolik proměnlivá, že je pojem „odpověď na podnět“ bezcenná. Odpověď nelze s ohledem na náhodný výsledek předvídat.
Vědecký pojem svobodné vůle/volního rozhodování se tím na rozdíl od metafyzického pojmu svobodné vůle pohybuje mezi krajnostmi naprostá náhoda/naprostá nutnost, čistý indeterminismus/tvrdý determinismus. Pravděpodobnost neboli stochasticita je součástí skutečnosti. Vědecký pojem svobodné vůle je kvantitativní, nikoli kvalitativní pojem. Otázka není správně položená, ptáme-li se: „Máme nebo nemáme svobodnou vůli?“ Správně položená otázka zní: „Kolik svobodné vůle jsme měli, máme nebo budeme mít?“ (7, 14, 15).
Současný americký filozof John Searle popsal svobodnou vůli jako přesvědčení, že „jsme se často mohli chovat jinak, než jsme se ve skutečnosti chovali“. Pijavky, švábi i drosofily se dokáží chovat ve stejném prostředí odlišně. Tento druh svobody je nutnou, nikoli však postačující složkou svobodné vůle.
Náhodné proměny neboli stochasticita chování jsou samy o sobě jen první složkou. Následuje rozhodnutí o tom, kterou z nabízených možností živočich nebo člověk volí. První krok je „stochastický“, druhý krok je „volní“, což odpovídá dvousložkovému modelu svobodné vůle navrženému jako prvním Williamem Jamesem a prosazovaném později mnoha dalšími vědci i filozofy, například Poincarém, Comptonem, Popperem, Dennetttem, Kanem, Fisherem, Melem, Kosslynem, Doylem, Kochem a Heisenbergem.



MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Thomayerova nemocnice
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Sources
1. Addis, D.R., Wong, A.T., Schacter, D.I. Remembering the past and imagining the future: common and distinct neural substrates during event construction and elaboration. Neuropsychologia, 2007, 45, p. 1363–1377.
2. Akkal, D., Dum, R.P., Strick, P.L. Supplementary motor area and presupplementary motor area: targets of basal ganglia and cerebellar output. J Neurosci, 2007, 27, p. 10659–10673.
3. Banks, P.W., Isham, E.A. We infer rather than perceive the moment we decided to act. Psychol Sci, 2009, 20, p. 17–21.
4. Brass, M., Haggard, P. To do or not to do: the neural signature of self control. J Neurosci, 2007, 27, p. 9141–9145.
5. Brass, M., Haggard, P. The hidden side of intentional action: the role of the anterior insular cortex. Brain Struct Funct, 2010, 214, p. 603–610.
6. Brembs, B. The importance of being active. J Neurogenet, 2008, 23, p. 120–126.
7. Brembs, B. Toward scientific concept of free will as a biological trait: spontaneous actions and decision-making in invertebrates. Proc R Soc B, 2011, 278, p. 930–939.
8. Buckner, R., Andrew-Hanna, J., Schacter, D. The brain´s default network: anatomy, function and relevance to disease. Ann NY Acad Sci, 2008, 1124, p. 1–38.
9. Custers, R., Aarts, H. The unconscious will: how the pursuit of goals operates outside of conscious awareness. Science, 2010, 329, p. 47–50.
10. Dobzhansky, T. Nothing in biology makes sense except in the light of evolution. Am Biol Teach, 1973, 35, p. 125–129. Dostupné též na: http://people.delphiforums.com/lordorman/light.htm.
11. Dennett, D. Consciousness explained. Boston: Little Brown, 1991.
12. Dennett, D., Kinsbourne, M. Time and the observer. Behav Brain Sci, 1992, 15, p. 183–274.
13. Domenici, P., Booth, D., Blagburn, J.M., et al. Cockroaches keep predators guessing by using preferred escapes trajectories. Curr Biol, 2008, 18, p. 1792–1796.
14. Doyle, R.O. Free will: it´s normal biological trait, not a gift or a mystery. Nature, 2009, 459, p. 1052.
15. Doyle, B. Jamesian free will. The two stage-model of William James. William James Studies 2010; 5, p. 1-28 [on-line]. Dostupné na: http://williamjamesstudies.org/5.1/doyle.pdf.
16. Fair, D.A., Cohen, A.L., Dosenbach, N.U.C., et al. The maturing architecture of the brain´s default network. Proc Natl Acad Sci USA, 2008, 105, p. 4028–4032.
17. Wikipedia. Free will [on-line]. Dostupné na: http://en.wikipedia.org/wiki/Free_will.
18. Friesen, W.O., Kristan, W.B. Leech locomotion: swimming, crawling, and decision. Curr Opin Neurobiol, 2007, 17, p. 704–711.
19. Haggard, P, Eimer, M. On the relation between brain potentials and the awareness of voluntary movement. Exp Brain Res, 1999, 126, p. 128–133.
20. Haggard, P. Human volition: towards a neuroscience of will. Nat Rev Neurosci, 2008, 9, p. 934–946.
21. Hall, L., Johansson, P., Tärning, B., et al. Magic at the marketplace: choice blindness for the taste of jam nad the smell of tea. Cognition, 2010, 117, p. 54–61.
22. Hallettt, M. Volitional control of movement: the fysiology of free will. Clin Neurophysiol, 2007, 118, p. 1179–1192.
23. Heisenberg, M. Is free will an illusion? Nature, 2009, 459, p. 164–165.
24. Hume, D. A treatise on human nature (1739). Dostupné na http://oll.libertyfund.org/index.php? option=com_staticxt&staticfile=show.php%3Ftitle=342&Itemid=28.
25. Koukolík, F. Evoluce a evoluční teorie pro lékaře. X. Sebeuvědomování. Prakt. lék., 2010, 90, s. 571–576.
26. Koukolík, F. Funkční systémy lidského mozku. Třetí vydání. Praha: Galén, 2012.
27. Kristan, W.B. Neuronal desicion-making circuits. Curr Biol, 2008, 18, R929–R932.
28. Kühn, S., Brass, M. Retrospective construction of the judgement of free choice. Conscious Cogn, 2009, 18, p. 12–21.
29. Leary, M.R., Buttermore, N.R. The evolution of human Self: tracing the natural history of self-awareness. J Theory Soc Behav, 2003, 33, p. 365–404.
30. Li C, S.R., Yan, P., Berquist, K.L., et al. Greater activation of the „default“ brain regions precede stop signal errors. NeuroImage, 2007, 38, p. 640–648.
31. Libet, B., Gleason, C.A., Wright, W., et al. Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential). The unconscious initiation of freely voluntary act. Brain, 1983, 106, p. 623–642.
32. London, M., Roth, A., Beeren, L., et al. Sensitivity to perturbation in vivo implies high noise and suggest rate coding in cortex. Nature, 2010, 466, p. 123–127.
33. Mason, M.F., Bortin, M.I., Van Horn, J.D., et al. Wandering minds: the default network and stimulus-independent thought. Science, 2007, 315, p. 393–395.
34. Matsuhashi, M., Hallett, M. The timing of the conscious intention to move. Eur J Neurosci, 2008, 28, p. 2344–2351.
35. Nachev, P., Rees, G., Parton, A, et al. Volition and conflict in human medial frontal cortex. Curr Biol, 2005, 15, p. 122–128.
36. Roskies, A. Neuroscientific challenges to free will and responsibity. Trends Cogn Sci, 2006, 10, p. 420–423.
37. Soon, Ch.S, Brass, M., Heinze, H.-J., et al. Unconscious determinants of free decisions in the human brain. Nat Neurosci, 2008, 11, p. 543–545.
38. Taylor, J.L., McCloskey, D.I. Selection of motor responses on the basis of unperceived stimuli. Exp Brain Res, 1996, 110, p. 62–66.
39. Wegner, D.M. The illusion of conscious will. Cambridge Massachusetts: MIT Press, 2002.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2012 Issue 4
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
- Memantine Eases Daily Life for Patients and Caregivers
Most read in this issue
- Astma a výživa: stravovací doporučení pro prevenci a léčbu astmatu
- Varianty lidských chromozomů a jejich význam z pohledu klinické genetiky
- Jaké je v České republice riziko onemocnění legionelózou?
- Význam histologické verifikace metastáz tumorů