
Proč se dráhy kříží? Základní principy uspořádání mozku obratlovců
Why do the nerve tracts decussate? Basic principles of the vertebrate brain organization
The fact that some brain tracts decussate accompanies medical students and neurologists and neurosurgeons during the whole period of their study, as well as in their work “careers”. We take the decussation of the pyramidal tract, anterolateral, lemniscal systems and visual pathways for granted and we describe contralateral hemipareses, hemiplegia, including alternating ones, Brown-Séquard’s spinal cord hemisyndrome and homonymous hemianopsia. We understand the central lesion of the facial nerve, presenting only with contralateral paresis of mouth muscles while the ability to close the eye is preserved. The other tracts which cross and in which we do not often realize this are for example the tracts of the dentate-rubro-olivary (Guillain-Mollaret’s) triangle, lemniscus lateralis (corpus trapezoideum) conducting hearing, tractus pontocerebellaris, and tractus dentatothalamicus (decussatio pedunculi cerebellaris superioris). The trochlear nerve, tectospinal tract (decussatio tegmenti dorsalis) and rubrospinal tract (decussation tegmenti ventralis) decussate in the mesencephalon, reticulospinal tracts are both crossed and also uncrossed and on the contrary the crossing of the interstitiospinal tract from the ncl. of Cajal and vestibulospinal tract from the ncl. of. Dieters are not present. We questioned this finding, so we conducted the review of the pertinent literature concerning the theories why the tracts cross, we added the mnemotechnic predator theory of the silur sea, and we concentrated on the phylogenetic differences in the architecture of vertebrate brains, with the help of dissection of brain cadavers from rabbit (Oryctolagus cuniculus), duck (Anas platyrhynchos domesticus) and carp (Cyprinus carpio). Based on our results, we are not able to answer all of the above-mentioned and further related questions, hence we present them to the reader in the form of a minimonography.
Keywords:
brain tract – comparative anatomy – Nervous system – corticospinal tract – optic chiasm – evolution
Authors:
R. Bartoš *1,2; D. Ospalík 3; A. Hejčl 1; A. Malucelli 1; M. Sameš 1; V. Němcová *2
Authors‘ workplace:
Veronika Němcová a Robert Bartoš, se na minimonografii podíleli stejným, dílem.
*; Neurochirurgická klinika UJEP, Masarykova, nemocnice, KZ a. s., Ústí nad, Labem
1; Anatomický ústav 1. LF UK, Praha
2; Neurologické oddělení, Masarykova, nemocnice, KZ a. s., Ústí nad Labem
3
Published in:
Cesk Slov Neurol N 2021; 84/117(4): 316-328
Category:
Minimonography
doi:
https://doi.org/10.48095/cccsnn2021316
Overview
Fakt křížení některých mozkových drah provází mediky a posléze neurology a neurochirurgy po celou dobu jejich studia, resp. po dobu jejich pracovní „dráhy“. Křížení pyramidové dráhy, anterolaterálního, lemniskálního systému a zrakové dráhy bereme jako samozřejmost a popisujeme kontralaterální hemiparézy, hemiplegie, vč. těch alternujících, Brown-Séquardův míšní hemisyndrom a homonymní hemianopsii. Chápeme centrální lézi lícního nervu projevující se pouze kontralaterální parézou koutku úst při zachování schopnosti zavřít oko. Dalšími drahami, které se kříží a u kterých si to často ani neuvědomujeme, jsou např. dráhy dentato-rubro-olivárního (Guillain-Mollaretova) trojúhelníku, lemiscus lateralis (corpus trapezoideum) vedoucí sluch, tractus pontocerebellaris a tractus dentatothalamicus (decussatio pedunculi cerebellaris superioris). Nervus trochlearis, dráha tektospinální (decussatio tegmenti dorsalis) a rubrospinální (decussation tegmenti ventralis) se kříží v mezencefalu, reticulospinální dráhy jsou zkřížené i nezkřížené, a naopak nedochází ke křížení dráhy interstitiospinální z Cajalova jádra a vestibulospinální z Dietersova jádra. Zkusili jsme se nad tímto zamyslet, provedli rešerši dostupné literatury ohledně teorií důvodů křížení drah, přidali mnemotechnickou predátorskou teorii silurského moře a zaměřili se na fylogenetické rozdíly ve stavbě mozku obratlovců disekcí kadaverózního mozku králíka (Oryctolagus cuniculus), kachny (Anas platyrhynchos domesticus) a kapra (Cyprinus carpio). Výsledky, byť nemůžeme odpovědět na všechny výše uvedené otázky a další otázky z nich vyvstávající, laskavému čtenáři předkládáme formou minimonografie.
Klíčová slova:
mozková dráha – srovnávací anatomie – nervový systém – kortikospinální dráha – chisama opticum – evoluce
Křížení pyramidové dráhy, anterolaterálního, lemniskálního systému a zrakové dráhy bereme v pozici klinických lékařů jako samozřejmost a popisujeme kontralaterální hemiparézy, hemiplegie, vč. těch alternujících, Brown-Séquardův míšní hemisyndrom a homonymní hemianopsii. Chápeme centrální lézi lícního nervu projevující se pouze kontralaterální parézou koutku úst při zachování schopnosti zavřít oko. Otázka důvodů a příčin zdánlivě nesmyslného křížení mnohých drah ovšem fascinuje neurovědce již dlouhou dobu. Je možné, že to ani nebyl neurovědec, kdo si této skutečnosti všimnul jako první. Traumatické krvácení do mozku při násilné a bojovné povaze člověka způsobilo postižení hybnosti na druhé straně než trauma mozku a již nějaký vnímavý Australophitecus africanus před 2–3 miliony let nebo vůdčí hominid (Ten, který hledí na Měsíc) v Clarcově Vesmírné odyseji (1968) mohli mít podobnou zkušenost – minimálně si to mohli prožít na vlastní kůži. Abychom se však dostali od spekulací k „seriózní vědě“ – tento fakt popsal Hippokrates (460–370 před Kristem). Všiml si totiž, že pokud má pacient (asi hoplít zraněný během peloponéské války) úraz na jedné straně hlavy, křeče se objevují na druhostranných končetinách. Až o 500 let později se ke stejnému jevu vrací Aretaeus z Cappadoccie (1.–2. století našeho letopočtu) ve spise „Morborum acutorum, de curatione apoplexiae“. Na základě svých pozorování, že pokud je sečná rána na jedné straně krční páteře, ochrnutí je na stejnostranných končetinách, dedukoval pravděpodobnou etáž křížení motorické dráhy. Až v roce 1710 François Pourfour du Petit, vojenský lékař armády Ludvíka XIV., ve svém pojednání „Lettres d’un medecin des hôpitaux du roi a un autre medecin de ses amis“ na základě anatomické disekce lokalizoval křížení pyramidové dráhy do oblasti prodloužené míchy. V roce 1810 Franz Joseph Gall (1758–1828) se svým žákem Johannem Spűrzheimem vypreparovali pyramidovou dráhu z dekuzace do oblasti kortexu, čímž potvrdili kontinuitu dráhy. Dalším Gallovým vědeckým počinem bylo geniální rozlišení funkce šedé a bílé hmoty mozkové, naopak typickým Cimrmanovým krokem stranou na pozice Bohlenova vulgárního materialismu bylo zavedení frenologie – oboru zkoumajícího souvislosti duševních schopností a charakterových rysů ze stavby lebky. Pro odvození duševních schopností frenologové používali nejen celkovou velikost a tvar lebky, ale především její povrchový reliéf. Pohmatem vám tedy mohli vybrat životního partnera nebo vás odmítnout jako uchazeče o zaměstnání.
Pokud jde o důvod křížení kortikospinální, spinotalamické, lemniskální a zrakové dráhy, první a zatím s určitými výhradami stále platnou hypotézu postuloval v roce 1897 slavný španělský neurovědec a histopatolog Santiago Ramón y Cajal (1852–1934). Zajímavostí je, že byl současně výtečným malířem a jeho obrázky neuronů a gliových buněk nás stále uchvacují. Objevil po něm pojmenované Cajalovy horizontální buňky, což jsou interneurony v neokortexu. V mládí se dokonce chtěl stát malířem, nicméně jiný umělec, malíř pouťových atrakcí, řekl jeho otci: „Ten chlapec nemá talent“. Musel tedy jít na medicínu a v roce 1906 společně s Camillem Golgim obdržel Nobelovu cenu.
V principu lze vysvětlit, že jeho hlavní myšlenkou bylo to, že obraz zevního světa na sítnici je obrácený čočkou (lens crystallina). V případě monokulárního vidění je tak obraz světa na sítnici obrácený, a ještě navíc diskontinuální (obr. 1), v případě binokulárního vidění „pouze“ obrácený. Křížení chiasmatu obraz opět složí dohromady, ovšem obraz v mozkových hemisférách zůstává prostorově obrácený. Křížení senzitivních a motorických drah nám umožní reálné objekty vnímat a reagovat na ně tak, jak jsou v prostoru skutečně umístěny [1–3]. Lépe nám tuto teorii pomůžou pochopit nákresy volně převzaté z původní Cajalovy práce [4] (obr. 1, 2). Taktéž situaci může přiblížit náš příklad s jednoduchým 4písmenným napsaným slovem, např. velký nápis KOLO, kdy si můžeme teoreticky představit, že na retině u očí, které nemají překryv zorných polí (levé vidí písmena KO / pravé vidí písmena LO), existuje jako OK/OL, následně tedy musí dojít ke zkřížení této retinální informace, aby v mozku byla sice obrácená, ale kontinuální (zprava doleva) tedy OLOK = pozpátku KOLO. V mozku tedy vznikne alespoň OLOK a další dráhy (motorické a senzitivní) se kříží, aby reagovaly a vnímaly KO nalevo a LO napravo (obr. 1).
Fig. 1. The diagram explaining the necessity of complete decussation
of the visual tract during monocular vision for preservation
of the continuity of the seen world, followed by crossing of
motor (M) and sensitive (S) tracts, enabling reaction to the real
position of the seen subject in space (in this case arrow apex).
Drawn by Monika Němcová.
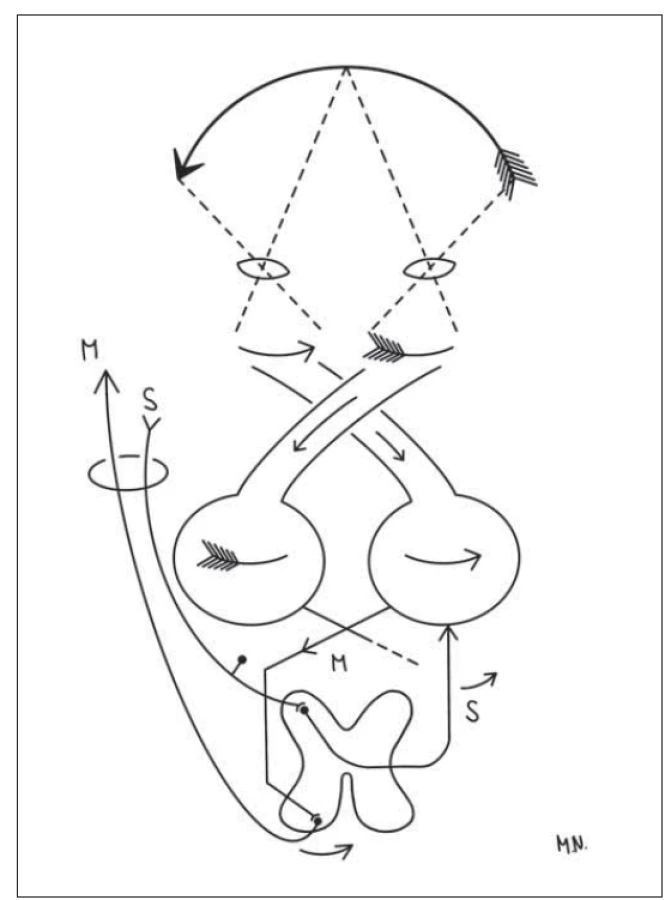
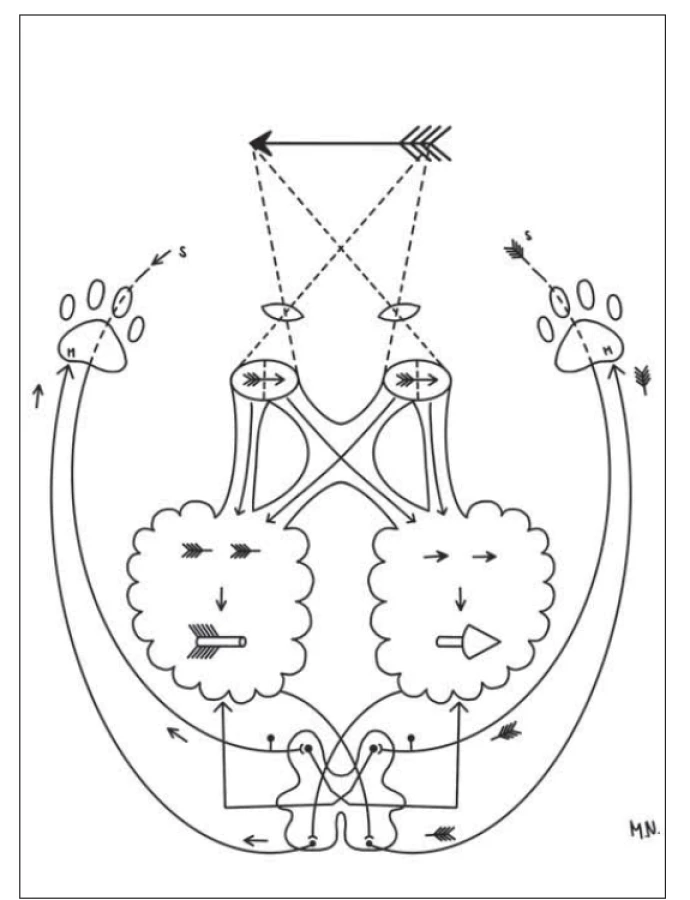
Fig. 2. The diagram explaining the partial crossing of the visual
tract in case of a binocularly seen object, its spatial synthesis in
the brain and crossing of the motor tract (M) in the medulla
oblongata and of the anterolateral anociceptive system (S) segmentally
in the spinal cord, again providing the correct spatial
determination of the seen object. Drawn by Monika Němcová.
Teorií, která není založena na zrcadlovém zobrazení viděného světa na sítnici, je teorie somatického twistu, kterou popsal v roce 2013 Marcel Kinsbourne [5]. U prvoústých (Protostomata), což jsou bezobratlí živočichové jako např. členovci, měkkýši a kroužkovci, je nervová soustava ventrálně od srdce i střev a až u druhoústých, kteří tvoří velice širokou skupinu živočichů – od ostnokožců (Echinodermata) po strunatce (Chordata), dojde k rotaci o 180° a řitní otvor se přesouvá na stejnou stranu těla jako ústní otvor, tedy ventrálně (obr. 3). Přesun CNS dorzálně s překřížením drah je jen „vedlejším produktem“ této rotace. Možná ale proto vzniká chorda dorzalis, aby CNS na ohrožené zádové straně chránila.
Fig. 3. (A) The scheme of the somatic twist in Deuterostomia, generating a 180º crossing of the nerve tracts. (B) An example of Protostomia:
azure damselfly (Coenagrion puella). (C) An example of Deuterostomia: nightingale bluethroat (Luscinia svecica). Both photos
and drawing: Antonín Cettl.

O něco složitěji tento somatický twist rozpracovali Marc de Lussanet a Jan Osse rok před ním [6]. Jejich twist je dvoufázový a o 90° proti sobě, výsledkem je tedy opět 180° rotace. Přinejmenším tak to kolegovi Kinsbournovi v roce 2015 vysvětlili [7]. Principem jejich teorie je přiložení levé strany embrya ke žloutkovému váčku (yolk sac), který u strunatců od ryb výše slouží k výživě zárodku. Aby mohl být žloutkový váček využíván, musí dojít k rotaci hlavové části embrya o 90° ve směru hodinových ručiček, zatímco zbytek těla kompenzatorně rotuje o 90° proti směru hodinových ručiček, čímž dochází ke křížení drah v úrovni chiasmatu a pod ním. Teorie somatického twistu však dle našeho názoru nevysvětluje, proč se spinotalamická dráha (tepelné a algické čití), přední spinocerebelární (propriocepce z dolní poloviny trupu a dolních končetin) a přední kortikospinální dráha (motorika trupu a šíje) kříží v míšním segmentu a jiné v různých částech mozkového kmene: laterální kortikospinální, část kortikonukleárních, rubrospinální, tektospinální dráha, dráha zadních provazců, pontocerebelární, olivocerebelární, cerebellorubrální a cerebelotalamická dráha. A proč se vestibulospinální a zadní spinocerebelární dráhy a část retikulospinálních ani kortikonukleárních drah vůbec nekříží? Naopak velice dobře vysvětluje fakt, proč jsou nervové dráhy nejkomplikovanějších prvoústých, žaludovců (Enteropneusta), nezkřížené a u nejprimitivnějších druhoústých (sliznatky a mihule) zkřížené, byť makroskopicky i způsobem života jsou si tito vodní živočichové velice podobní [8].
Existují i teorie matematicky nevzdělanému laikovi velice obtížně pochopitelné, využívající modelování 3D prostorových sítí. Uspořádání živých organizmů je jistě třídimenzionální, což potvrzuje větší anti-chybovou stabilitu přenosu dat zkřížených systémů oproti nezkříženým. Tato výhoda je silnější se zvyšujícím se počtem recipročních bodů ve 3D datové síti [9].
Během vývoje mozku je nezbytné, aby rostoucí axony projekčních a komisurálních neuronů (např. pyramidové dráhy nebo corpus callosum) v určitém místě překročily střední čáru a pokračovaly kontralaterálně a aby se nevracely zpět na původní stranu. K tomu slouží celá řada molekulárních mechanizmů, z kterých ještě mnohé neznáme. Jsou to geny kódující membránové receptory na povrchu axonů i ligandy, které se na ně při střední čáře navazují. Určité látky lákají rostoucí axony ke střední čáře (chemoatraktanty) a jiné je po překonání střední čáry odrazují od toho, aby se k ní znovu vracely (repelenty). V míše během vývoje fungují jako atraktanty Netriny produkované spodinovou ploténkou, ty se váží na receptor DCC (deleted in colorectal cancer) a jeho příbuzné (Neo, down syndrome cell adhesion molecule [DSCAM]). Dalšími atraktanty jsou sonic hedgehog (SHH) a vascular endotheliar growth factor (VEGF). Ve stropové ploténce tvořené BMPs (bone morphogenetic proteins) odpuzují rostoucí axony. Proto se commissura anterior alba vytváří ventrálně. Po překročení střední čáry pokračují axony kontralaterálně a nesmí se znovu křížit. To zabezpečují Slit proteiny, produkované gliemy spodinové ploténky, které se váží na Robo (Roundaboud) receptory na rostoucím axonu. Robo 3 má dvě varianty, 3.1 a 3.2. Než překročí axon střední čáru, je na jeho povrchu zvýšená exprese Robo 3.1 receptorů, což podporuje jejich růst ke střední čáře. Po překročení střední čáry je v membráně axonů více Robo 3.2 receptorů, jejichž aktivace pomáhá axonům mířit od střední čáry. Při mutacích, kde chybí Robo 3 receptor, nedochází ke zkřížení rostoucích axonů v míše. Robo 1 a Robo 2 receptory jsou na rostoucích axonech před překřížením zastoupeny minimálně, po překročení střední čáry se jejich exprese zvýší a spolupracují s aktivovanými receptory Robo 3 [10–12].
Zajímavostí evoluce jsou některé syndromy, např. Klippel-Feilův syndrom, kdy pacienti mají krátký krk s fúzemi krčních obratlů a nízko položenou vlasovou linii. Zajímavější však je, že pyramidová dráha se u nich nekříží v úrovni prodloužené míchy, ale segmentálně v míše, tudíž u 75 % pacientů při volním pohybu vznikají „zrcadlové pohyby“ druhostrannými končetinami [3,13]. Dalším syndromem je na chromozom X-vázaný Kallmannův syndrom, který je spojený s anosmií a hypogonadotropním hypogonadismem. Popsal jej Aureliano Maestre de San Juan (1828–1890), a jelikož je svět malý, byl právě tento vědec Cajalovým profesorem histologie. U 85 % pacientů s tímto vzácným syndromem dochází při magnetické transkraniální stimulaci k současné ko - aktivaci končetin obou stran, zdá se tedy, že podstatná část kortikospinálního traktu probíhá nezkříženě. Anatomický sekční korelát chybí, kromě defektního genu ANOS1 (dříve KAL-1) se dle recentních prací může jednat i poruchu genů Robo1/2 [3,14–16]. U lidí vzácný syndrom poruchy horizontálního pohledu spojený se skoliózou (horizontal gaze palsy with progressive scoliosis; HGPPS) je způsoben mutací robo genu Robo3. V mozkovém kmeni a míše při něm nedochází ke zkřížení dráhy spojující jádro nervus abducens s jádrem nervus oculomotorius, proto je zde obrna horizontálního pohledu. Vertikální oční pohyby jsou normální. Při vyšetření MR byla prokázána hypoplazie nucleus abducens (není patrný colliculus facialis) a změny tvaru pontu a oblongáty spojené s úbytkem bílé hmoty. Evokované potenciály lidí s tímto syndromem prokazují, že mají nezkříženou jak pyramidovou dráhu, tak i lemniskální systém. Přesto se tito lidé normálně pohybují a mají normální dotykové čití i propriocepci. Těžká skolióza je zřejmě neurogenní, z úbytku retikulospinálních drah odpovědných za svalový tonus [17,18]. U myší s mutací genů Rig-1/Robo 3 se nekříží vlákna z precerebelárních jader kmene vč. dolní olivy [19].
Potud se jednalo o výčet některých teorií a mechanizmů křížení drah. Kromě výše uvedených a do jisté míry pro „život a lidstvo“ optimističtějších teorií existují predátorské teorie vysvětlující křížení drah na jednoduchém a utilistaristickém modelu (sežrat, ale přitom nebýt sežrán). Např. Sarnat a Netzky v roce 1974 [20] pozorovali defenzivní kroutivý reflex u kopinatce plžovitého (Branchiostoma lanceolatum), což je primitivní strunatec trávící většinu života zahrabaný v písku. Tento reflex vedoucí přímou zkříženou dráhou mu umožňuje rychlou ochranu napadené strany těla okamžitou kontrakcí svalstva druhé strany. Tyto teorie nám mohou být sympatické asi jen z toho důvodu, že nám mohou pomoci mnemotechnicky ulehčit představu a zapamatování si komplikovaného křížení motorických a senzitivních drah.
Naše teorie napadla prvního autora nezávisle na výše uvedené citaci během pádlování na rychlostním kajaku (pádlo = ploutev). Představte si, že plavete v pravěkém moři před 420 miliony let a jste velký primitivní predátor (obr. 4). Vidíte něco (co chcete sežrat) na vaší pravé straně. Zmiňme se nejprve o oku, komorové oko s čočkou není samozřejmostí, byť se zřejmě vyvinulo nezávisle třikrát – u obratlovců (Craniata), hlavonožců (Cephalopoda) a u medúz z třídy čtyřhranky (Cubozoa). Jako správný predátor máte již binokulární vidění a čočku, tedy oči vpředu a m. protractor lentis, a tak máte primárně zaostřeno na dálku. Zrakový vjem je přenesen do levé i do pravé hemiféry a je rychlejší zabrat levou přední ploutví, a tím zahnout doprava. Pokud jde nervový impuls přímo zkříženě laterální kortikospinální (pyramidovou) dráhou, je to rychlejší. Máte výhodu, pokud je to i bez interneuronu. Teď si naopak představte, že jste ta malá kousnutá primitivní rybka nebo trilobit. „Ale co to? Něco se teď dotklo mé levé strany.“ – to je propriocepce. Impulz jde v medulle oblongatě cestou křížících se zadních provazců (lemniskální systém) do pravé hemisféry – tudíž pokud mám čas, pokusím se uprchnout doprava pohybem levé přední ploutve, což je opět přímý a zkřížený pyramidový systém. „Ale ne, ta bestie“ byla moc rychlá a zakousla se do mojí levé poloviny trupu, poničila ji a ploutev nefunguje – pokud tedy chci cítit bolest, která je nutná pro následnou záchrannou akci, vzruch musí jít „nahoru“, segmentálně se křížícími vlákny anterolaterálního systému (spinotalamického traktu). Pokud budu chtít alespoň „mrskat“ ocasem, abych se pokusil uprchnout, může do něj vzruch jít pravou polovinou těla, nezkříženým anteriorním kortikospinálním traktem, který se kříží až segmentálně pod místem destrukce na straně útoku.
Fig. 4. Eurypterus hunts the trilobite
(Přídolí period, silur, aprox. 420 million
years ago). Drawn by Antonín Cettl.
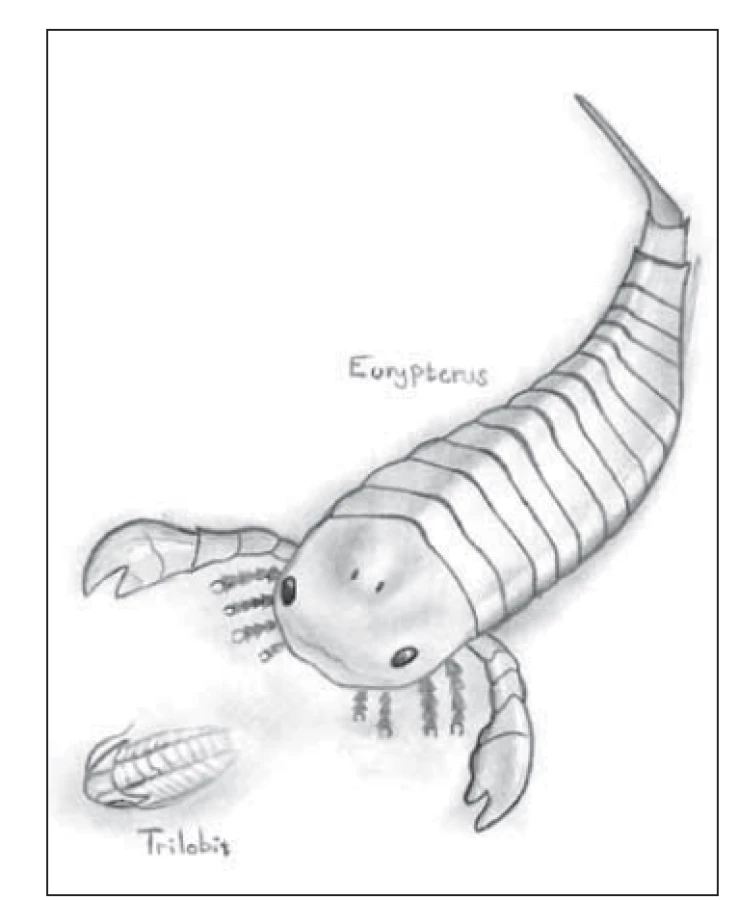
Tato teorie však naráží na několik „detailů“. Trilobit i jeho pronásledovatel Eurypterus jsou členovci, tedy prvoústí s žebříčkovitou nervovou soustavou pod trávicí trubicí. Fungují tedy jinak. V naší úvaze však nemůžete být ani rybou ani jiným „nesavcem“, jelikož ti nemají kortikospinální dráhu (pallio-spinální). U ptáků vede zkřížená dráha z telencefala jen do krční míchy. Ježci, damani a krtci nemají pyramidovou dráhu zkříženou [21]. I motorika králíka (Oryctolagus cuniculus), výborného běžce, kličkaře a skokana, je dominantně řízena v mezencefalu zkřížené rubrospinální dráze. Má však výborně vyvinutou kortikobulbární dráhu zajišťující výbornou hybnost obličejového svalstva pro jemný pohyb pysky a nasávání vzduchu do chřípí.
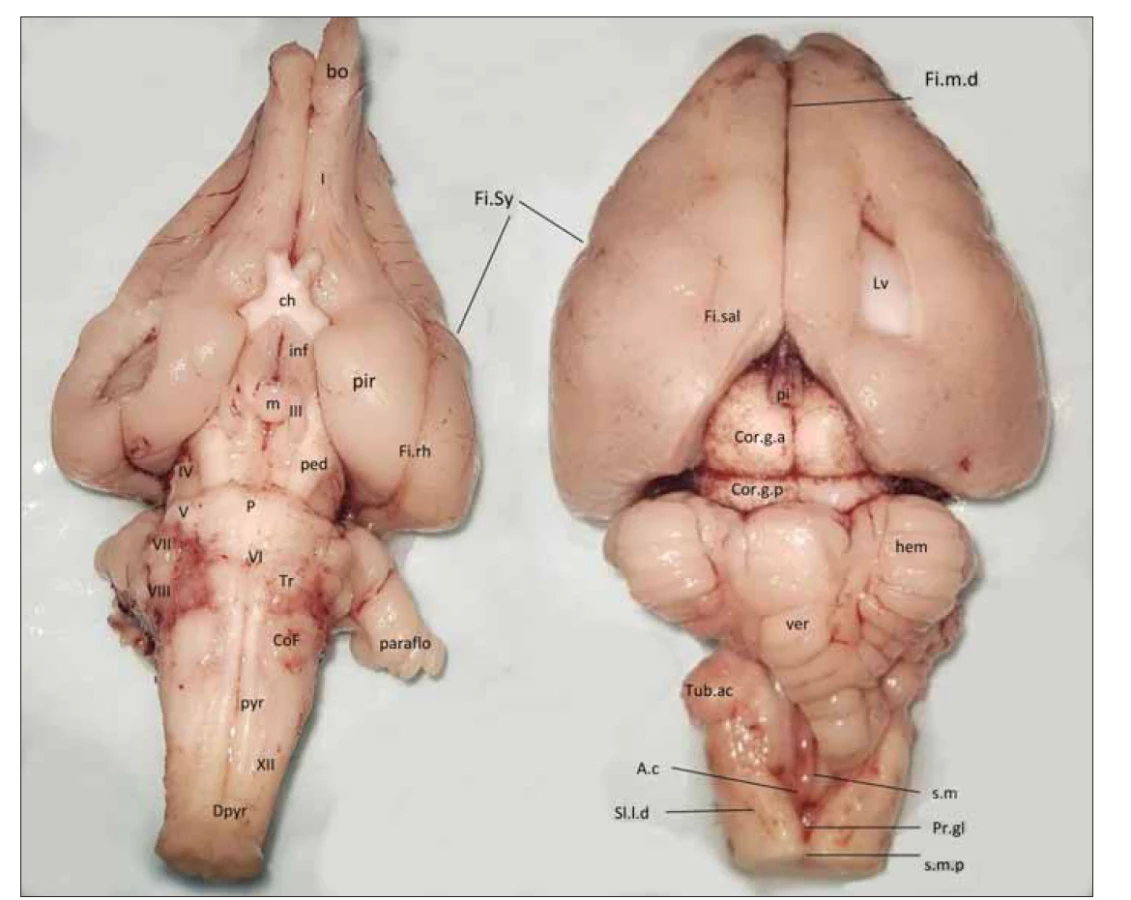
Preparát mozku králíka nám zobrazuje obr. 5. Zde si povšimněme lissencefalní mozkové kůry s pouze naznačenými fissurami a masivním olfaktorním bulbem, přímo navazujícím na piriformní kortex. Lze předpokládat, že celý mediotemporální lalok je čichovým mozkem. Pod okcipitálními laloky vidíme velké čtverohrbolí, což je zraková, resp. sluchová kmenová oblast. Králík má dobrý sluch, proto se dorzálně při výstupu vestibulokochleárního nervu vyklenuje velké tuberculum acusticum (sluchové jádro) a na ventrální ploše kmene je povrchově uložené corpus trapezoideum (ncl. olivaris superior a ncl. corporis trapezoidei a jejich spoje). Jak jsme uvedli, transverzální pontinní vlákna jsou zatím málo vyvinutá, takže můžeme sledovat takřka celý průběh pyramidové dráhy ve kmeni od crura cerebri po křížení pyramidové dráhy. Naopak uspořádání a odstupy hlavových nervů jsou podobné, jako je pozorujeme u člověka. Hlavním rozdílem je kaudálně uložené, ventrálně vyklenuté jádro nervus facialis (tvoří colliculus facialis, který ovšem neodpovídá lidskému colliculus facialis dorzálně vyklenutému na spodině IV. komory). Králík patří mezi savce s laterálně postavenýma očima a má panoramatická zrakové pole, aby mohl dobře pozorovat blížící se lišku, většina vláken optických nervů se tedy musí křížit. Asi 10 % nezkřížených vláken míří hlavně do ncl. corporis geniculati dorsalis a do colliculus superior. Pouze předměty v úzkém zorném poli před sebou vidí binokulárně. V mozečku pozorujeme masivní vermis, relativně malé hemisféry mozečku a velký samostatný lobulus parafloccularis. Vermis a lobulus parafloccularis tvoří vestibulocerebellum, které králík nebo jeho příbuzný zajíc využívají při kličkovacích manévrech, kdy potřebuje perfektně analyzovat a zpracovat signály z rovnovážného ústrojí. My již tolik neskáčeme a právníci kličkují jenom mezi paragrafy, tudíž se paraflocullární lobulus zřejmě změnil na tonsillu a lobulus biventer. Vlastní flocullus máme velmi malý a skrytý pod objemnými hemisférami mozečku.
I – tracus olfactorius; III – n. oculomotorius; V – n. trigeminus; VI – n. abducens; VII – n. facialis; VIII – n. vestibulocochlearis; XII – n. hypoglossus;
A.c – ala cinerea (trigonum nervi vagi); bo – část bulbus olfactorius; ch – chiasma opticum; CoF – colliculus facialis; Cor.g.a – corpus geniculatum
anterius; Cor.g.p – corpus geniculatum posterius; Dpyr – decussatio pyramidum; Fi.m.d – fissura mediana dorsalis; Fi.rh – fissura rhinalis;
Fi.sal – fissura sagittalis lateralis; Fi.Sy – Sylvijská rýha; hem – mozečková hemisféra; Inf – infundibulum hypophysae; Lv – ventriculus lateralis;
m – corpus mamillare; P – pons Varoli; paraflo – paraflocculus; ped – pedunculus cerebri; pi – corpus pineale; pir – piriformní kůra; Pr.gl – promontorium
gliosum calami scriptorii; pyr – pyramis medullae oblongatae; s.l.d – sulcus lateralis dorsalis (posterolateralis); s.m – sulcus medianus;
s.m.p – sulcus medianus posterior; Tr – corpus trapezoideum; Tub.ac – tuberculum acusticum; ver – vermis cerebelli
Fig. 5. The brain of the rabbit (left – ventral side, right – dorsal side).
I – tracus olfactorius; III – n. oculomotorius; V – n. trigeminus; VI – n. abducens; VII – n. facialis; VIII – n. vestibulocochlearis; XII – n. hypoglossus;
A.c – ala cinerea (trigonum nervi vagi); bo – part of the bulbus olfactorius; ch – chiasma opticum; CoF – colliculus facialis; Cor.g.a – corpus geniculatum
anterius; Cor.g.p – corpus geniculatum posterius; Dpyr – decussatio pyramidum; Fi.m.d – fissura mediana dorsalis; Fi.rh – fissura
rhinalis; Fi.sal – fissura sagittalis lateralis; Fi.Sy – sulcus lateralis (Sylvii); hem – cerebellar hemisphere; Inf – infundibulum hypophysae; Lv – ventriculus
lateralis; m – corpus mamillare; P – pons Varoli; paraflo – paraflocculus; ped – pedunculus cerebri; pi – corpus pineale; pir – piriform
cortex; Pr.gl – promontorium gliosum calami scriptorii; pyr – pyramis medullae oblongatae; s.l.d – sulcus lateralis dorsalis (posterolateralis);
s.m – sulcus medianus; s.m.p – sulcus medianus posterior; Tr – corpus trapezoideum; Tub.ac – tuberculum acusticum; ver – vermis cerebelli
Pokud by měl čtenář pocit, že nám popis mozku králíka pomohl pochopit lidský mozek a že pokud půjdeme „evolučně níže“ podle Edingerovy teorie lineární evoluce [22], vše bude jednodušší, opak je pravdou. Mozek ptáků není překvapivě vůbec zjednodušenou verzí savčího mozku. Ptáci pocházejí ze společného předchůdce sauropsidní linie, ze které se vyvinuli společně s plazy (Reptilia). K oddělení sauropsidní linie došlo zhruba před 240 miliony let. Ariëns Kappers v roce 1936 v knize „The Comparative Anatomy of the Nervous System of Vertebrates Including Man“ [23] rozdělil telencefalon na pallium – oblast dorzálně od komor a subpallium – u savců septum a bazální ganglia. U ptáků tvoří typické vrstevnaté pallium jen bulbus olfactorius, archikortex (piriformní kůra a hippokampální formace) a úzká dorzolaterální kortikální oblast. Dorzálně od bazálních ganglií (archistriata – amygdala, paleostriatálního komplexu – globus pallidus) se nachází dorzální komorový hřeben (savčí neostriatum, tedy: putamen, ncl. caudatus a ncl. accumbens) a tzv. hyperstriatum, neboli Wulst, podélný hrbol v dorzální části hemisféry. Na základě pozorování inteligence ptáků, studia spojů a exprese genů v hemisféře ptáků, plazů a savců, Reiner et al v roce 2004 vytvořili zcela jinou nomenklaturu pro ptačí telencefalon [24]. Výše popsaný Wulst se nazývá hyperpallium, je lamelárně uspořádán a projikuje se do něj senzitivita cestou lemniskálního systému a dále zrakové informace, eferenty potom míří do míchy a kmene. Zbytek telencefala tvoří dorsal ventricular ridge (DVR), tedy dorzální komorový hřbet, který se z povrchu do hloubky dělí na mezopallium, nidopallium a arcopallium (obr. 6) [25]. Mezopallium a nidopallium jsou považovány za místa ptačího myšlení, učení a paměti. Mesopallium dorsale je obousměrně propojeno s Wulstem a mesopallium ventrale dostává signály z entopallia (zrak) a pole L (sluch) a po jejich zpracování vysílá signály do nidopallia a arcopallia. Někteří autoři považují mezopallium za 2. a 3. vrstvu neokortexu savců (asociační interneurony), jiní se domnívají že jsou mezopalllium a nidopallium homologní claustru a amygdale. Do vmezeřených částí nidopallia míří informace z trigeminu – rostrálně umístěný ncl. basorostralis pallii, zrakové dráhy (uprostřed ležící entopallium) a sluchové dráhy (vzadu ležící pole L). Do oblouku stočené arcopallium se dále dělí na kaudomediální část, která je homologní k amygdale, a anteriorní část homologní k senzomotorické oblasti neokortexu. Stavbou se DVR podobá bazálním gangliím, tak jak to popsal Kappers, ale typy neuronů, spoje a funkce odpovídají savčímu neokortexu. Laminární uspořádání má pouze Wulst, hyperpallium, od DVR oddělené na povrchu valleculou a uvnitř pomocí lamina frontalis superior. Neobvyklé uspořádání neuronů v hemisféře ptáků nesvědčí o nižší úrovni organizace jejich mozků oproti savcům (zde např. králíkovi). Je známo, že vrána novokaledonská (Corvus moneduloides) si dokáže vyrobit nástroj, a to i složením z několika součástí. V pokusech také dokáže využívat sofistikované triky založené na porozumění fyzikálním zákonům, např. do válce s vodou, ve které plave pamlsek, vhazuje další předměty, a přitom je schopna odhadnout hustotu i velikost vhazovaného předmětu. Je schopna pamlsek získat i manipulací s U-trubicí se spojenou hladinou, kdy tento ovšem plave v úzkém ramenu trubice a předměty musejí být vhazovány do širšího ramene, ve kterém pamlsek není. K tomu připočtěme schopnost opakovat a učit se slova u papoušků, schopnost geografické orientace tažných ptáků a můžeme konstatovat, že v některých ohledech se ptáci jistě vyrovnají primátům či kytovcům. Došlo pouze k tomu, že neurony s homologní funkcí jako u synapsidní (savčí) větve nemigrovaly dorzolaterálně do horního pallia, ale proliferovaly in situ a expandovaly do postranní komory.
Na dorzálním povrchu hemisféry je červený vrstevnatý Wulst (hyperpallium), do kterého projikuje lemniskální systém a zraková dráha. Eferenty
z něj míří do kmene a míchy. Vzadu za ním je fialový archicortex. Další části ptačího mozku už nejsou vrstevnaté. Pod Wulstem je zelené
mezopallium rozdělené přerušovanou čarou na dorzální mezopallium, obousměrně propojené s Wulstem, a ventrální mezopallium, které dostává
senzorické informace z entopallia (žlutě) a po zpracování je posílá do nidopallia (modře) a arcopallia (světle fialově). Kaudomediální část
arcopallia odpovídá amygdale a přední část senzorimotorické oblasti neokortexu. Entopallium vybíhá směrem do nidopallia (vmezeřené nidopallium)
a má vpředu senzitivní nucleus basorostralis pallii, uprostřed zrakovou oblast a vzadu sluchové pole L. Mezopallium, nidopallium
a entopallium se dohromady nazývají dorsal ventricular ridge (zadní komorový hřeben). V hloubce hemisféry jsou bazální ganglia (oranžově
striatum a světle oranžově pallidum). Šedě je mozkový kmen a mozeček. Primární senzitivní neurony ptačího mozku sedí v entopalliu (žlutě),
v nidopalliu (modře) a ve vmezeřeném hyperpalliu (růžově), kde jsou zrakové a somatosenzitivní oblasti. Hlavním korovým eferentem jsou
neurony arcopallia. Napříč popsanými oblastmi ptačího mozku prostupují funkční sloupce barevně ohraničené: modře senzitivita, červeně
motorika, zeleně zrak, fialově sluch; ve Wulstu jsou to zezadu dopředu sloupce pro noční vidění (ohraničený zeleně), denní vidění (žlutě) a somatosenzitivitu
(modře).
Crbl – mozeček; Hip – archicortex; Mes – mesencephalon; Rh – rhombencephalon; SSenz – somatosenzitivita; TeO – tectum opticum;
Th – thalamus
Fig 6. The diagram of the brain of the bird with color coded parts of the hemisphere and color bordered functional areas dividing the
hemisphere. According to [25], drawn by Monika Němcová.
On the dorsal surface of the hemisphere, there is the red laminar Wulst (hyperpallium), into which the lemniscal system and visual tract project.
Its eff erents project towards the brain stem and spinal cord. The violet archicortex is behind. Other parts of the bird brain are not laminar.
Under the Wulst, there is the green mesopallium, divided by the interrupted line to the dorsal mesopallium, which is bidirectionally connected
with the Wulst, and ventral mesopallium; this receives sensory input from the entopallium (yellow) and after its processing it sends
the information to the nidopallium (blue) and arcopallium (mauve). Caudomedial part of the arcopallium corresponds to the amygdala and
the anterior part of the sensomotor area of the neocortex. Entopallium plunges in the direction of the nidopallium (interposed nidopallium)
and anteriorly has a sensitive nucleus basorostralis palliii, and in the middle is the visual area and posteriorly auditory field L. The common
name for the mesopallium, nidopallium, and entopallium is the dorsal ventricular ridge. In the depth of the hemisphere, there are basal ganglia
(orange striatum and light orange pallidum). The gray color represents the brain stem and cerebellum. Primary sensitive neurons of the
bird brain lay in the entopallium (yellow), nidopallium (blue), and interposed nidopallium (pink), which are visual and somatosensitive areas.
The arcopallium neurons constitute the main cortical eff erent. Through the described areas of the bird brain, functional columns pervade,
which are color bordered: blue = sensitivity, red = motor activity, green = vision, violet = hearing; in the Wulst, these are from the posterior to
the anterior columns including night vision (green), daylight vision (yellow), and somatosensitivity (blue).
Crbl – cerebellum; Hip – archicortex; Mes – mesencephalon; Rh – rhombencephalon; SSenz – somatosensory; TeO – tectum opticum;
Th – thalamus
![Schéma ptačího mozku s barevně vyznačenými částmi hemisféry a s barevně ohraničenými funkčními oblastmi jdoucími napříč
částmi hemisféry. Podle [25], nakreslila Monika Němcová.<br>
Na dorzálním povrchu hemisféry je červený vrstevnatý Wulst (hyperpallium), do kterého projikuje lemniskální systém a zraková dráha. Eferenty
z něj míří do kmene a míchy. Vzadu za ním je fialový archicortex. Další části ptačího mozku už nejsou vrstevnaté. Pod Wulstem je zelené
mezopallium rozdělené přerušovanou čarou na dorzální mezopallium, obousměrně propojené s Wulstem, a ventrální mezopallium, které dostává
senzorické informace z entopallia (žlutě) a po zpracování je posílá do nidopallia (modře) a arcopallia (světle fialově). Kaudomediální část
arcopallia odpovídá amygdale a přední část senzorimotorické oblasti neokortexu. Entopallium vybíhá směrem do nidopallia (vmezeřené nidopallium)
a má vpředu senzitivní nucleus basorostralis pallii, uprostřed zrakovou oblast a vzadu sluchové pole L. Mezopallium, nidopallium
a entopallium se dohromady nazývají dorsal ventricular ridge (zadní komorový hřeben). V hloubce hemisféry jsou bazální ganglia (oranžově
striatum a světle oranžově pallidum). Šedě je mozkový kmen a mozeček. Primární senzitivní neurony ptačího mozku sedí v entopalliu (žlutě),
v nidopalliu (modře) a ve vmezeřeném hyperpalliu (růžově), kde jsou zrakové a somatosenzitivní oblasti. Hlavním korovým eferentem jsou
neurony arcopallia. Napříč popsanými oblastmi ptačího mozku prostupují funkční sloupce barevně ohraničené: modře senzitivita, červeně
motorika, zeleně zrak, fialově sluch; ve Wulstu jsou to zezadu dopředu sloupce pro noční vidění (ohraničený zeleně), denní vidění (žlutě) a somatosenzitivitu
(modře).<br>
Crbl – mozeček; Hip – archicortex; Mes – mesencephalon; Rh – rhombencephalon; SSenz – somatosenzitivita; TeO – tectum opticum;
Th – thalamus<br>
Fig 6. The diagram of the brain of the bird with color coded parts of the hemisphere and color bordered functional areas dividing the
hemisphere. According to [25], drawn by Monika Němcová.<br>
On the dorsal surface of the hemisphere, there is the red laminar Wulst (hyperpallium), into which the lemniscal system and visual tract project.
Its eff erents project towards the brain stem and spinal cord. The violet archicortex is behind. Other parts of the bird brain are not laminar.
Under the Wulst, there is the green mesopallium, divided by the interrupted line to the dorsal mesopallium, which is bidirectionally connected
with the Wulst, and ventral mesopallium; this receives sensory input from the entopallium (yellow) and after its processing it sends
the information to the nidopallium (blue) and arcopallium (mauve). Caudomedial part of the arcopallium corresponds to the amygdala and
the anterior part of the sensomotor area of the neocortex. Entopallium plunges in the direction of the nidopallium (interposed nidopallium)
and anteriorly has a sensitive nucleus basorostralis palliii, and in the middle is the visual area and posteriorly auditory field L. The common
name for the mesopallium, nidopallium, and entopallium is the dorsal ventricular ridge. In the depth of the hemisphere, there are basal ganglia
(orange striatum and light orange pallidum). The gray color represents the brain stem and cerebellum. Primary sensitive neurons of the
bird brain lay in the entopallium (yellow), nidopallium (blue), and interposed nidopallium (pink), which are visual and somatosensitive areas.
The arcopallium neurons constitute the main cortical eff erent. Through the described areas of the bird brain, functional columns pervade,
which are color bordered: blue = sensitivity, red = motor activity, green = vision, violet = hearing; in the Wulst, these are from the posterior to
the anterior columns including night vision (green), daylight vision (yellow), and somatosensitivity (blue).<br>
Crbl – cerebellum; Hip – archicortex; Mes – mesencephalon; Rh – rhombencephalon; SSenz – somatosensory; TeO – tectum opticum;
Th – thalamus](https://www.prelekara.sk/media/cache/resolve/media_object_image_large/media/image_pdf/565f5ea5a3bae428abe6e10d5c9c000f.png)
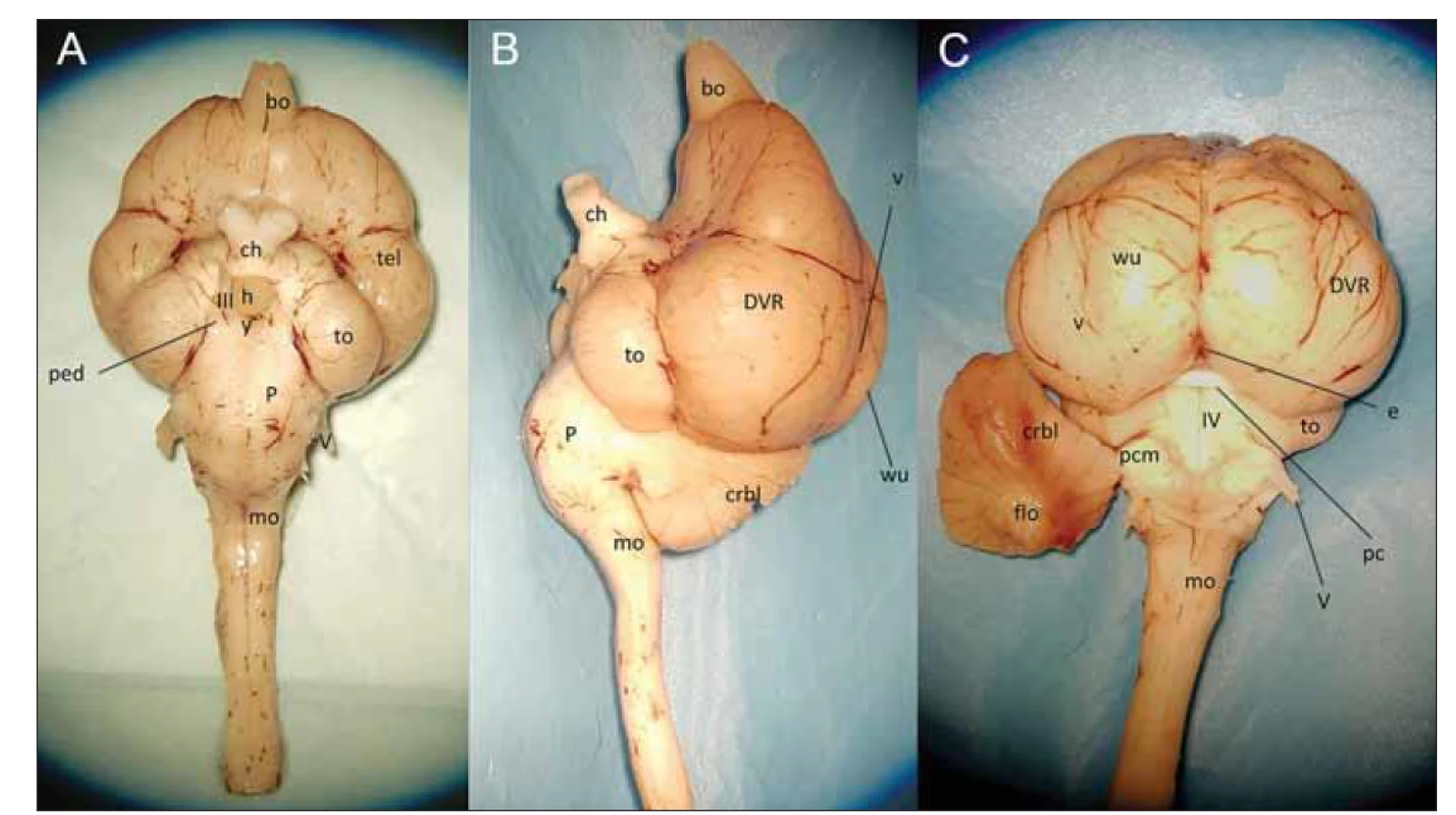
Mozek ptáků dokumentujeme preparátem kachny domácí (Anas platyrhynchos domesticus) na obr. 7. Na mozku kachny je stejně jako u ostatních ptáků nápadně velké tectum opticum. Má složitou vrstevnatou strukturu a převádí zrakové informace na motoriku kmene, integruje sluchové (dobře vyvinuté u sov), zrakové (u sokola) i taktilní signály (dobře vyvinutá senzitivita zobáku papoušků), posílá spoje zpět do retiny přes ncl. isthmi opticus a vysílá i ascendentní spoje. Hlavním ptačím smyslem bývá zrak na úkor čichu (výjimku představuje např. pták kiwi). Protože tectum opticum dostává 90 % projekce ze sítnice, je za hlavní zrakovou dráhu u ptáků považována dráha tektofugální: sítnice – tectum opticum – nucleus rotundus (odpovídá pulvinaru thalami u savců) – entopallium. V entopalliu se potom zpracovává informace o jasu, rozlišení, barvě, jednoduchých i složitých pohybech. Selektivní pozornost umožňuje propojení tectum opticum s nucleus isthmi a semilunaris. Druhou zrakovou dráhou je talamofugální: sítnice – ncl. opticus thalami (odpovídá savčímu corpus geniculatum laterale) – Wulst. Tato dráha je považována za homologní k savčí retino-geniculo-kortikální dráze končící ve striatické kůře V1. Hraje roli v prostorové orientaci, percepci pohybu a binokulárním vidění (dobře vyvinutá u sov). Ptáci dále mají akcesorní optický systém, který používají k analýze optického toku vzniklého vlastním pohybem a k tvorbě optokinetické odpovědi. Tvoří ho spojení mezi sítnicí a dvěma jádry v mozkovém kmeni (nucleus lentiformis mesencephali a nucleus of the basal optic root tectum opticum). Poslední zajímavou dráhou je spojení mezi tectum opticum, ncl. isthmo-opticus a retinou. Isthmo-optické jádro projikuje do sítnice k amakrinním buňkám a slouží k přepínání pozornosti a rychlému zaostření na blízko (sběr potravy) a do dálky (predátor). Toto jádro mají dobře vyvinutá kuřata, holubi, pěvci, datlové a kolibříci. Z ptačích smyslů kromě výborně sluchově vybavených sov stojí za zmínku i orientace tažných ptáků. Kromě orientačních bodů zachycených zrakem vnímají také magnetické pole země pomocí receptorů v horní části zobáku, odkud je informace pomocí nervus trigeminus vedena trigeminálním lemniskem do entopallia. Ztráta čichu pak vede u holubů k naprosté dezorientaci. Corpus callosum není u ptáků přítomno a nahrazují jej přední a zadní commissura. Mozeček je u ptáků dobře vyvinut, tvoří 12 % objemu mozku a je rozdělen na 10 hlavních folií, na sagitálním řezu se podobá savčímu mozečku. Laterálně prominující ouško se stejně jako u savců nazývá flocculus a tvoří se společně se zadní částí IX a X folia (u savců uvula a nodulus). Většinu mozečku tvoří spinocerebellum, za neocerebellum se považuje jen malá laterální část hemisfér bez folií. To, co vypadá jako velké vermis, je pravděpodobně ptačí mozečková hemisféra, protože ke svému vývoji nepotřebuje Fgf8. Vermis je výlučně savčí strukturou.
III – n. oculomotorius; IV – IV. komora; V – n. trigeminus; bo – bulbus olfactorius; ch – chiasma opticum; crbl – cerebellum; e – epiphysis; h –
hypothalamus; mo – medulla oblongata; p – pons Varoli; pc – commissura posterior; pcm – pedunculus cerebellomedullaris; ped – pedunculus
cerebri; pyr – pyramis medullae oblongatae; DVR – zadní komorový hřeben; to – tectum opticum; v – vallecula; wu – hyperpallium (hyperstriatum
– Wulst)
Fig. 7. The brain of the duck – (A) ventral view; (B) lateral view; (C) posterior view.
III – n. oculomotorius; IV – ventriculus quartus; V – n. trigeminus; bo – bulbus olfactorius; ch – chiasma opticum; crbl – cerebellum; e – epiphysis;
h – hypothalamus; mo – medulla oblongata; p – pons Varoli; pc – commissura posterior; pcm – pedunculus cerebellomedullaris; ped
– pedunculus cerebri; pyr – pyramis medullae oblongatae; DVR – dorsal ventricular ridge; to – tectum opticum; v – vallecula; wu – hyperpallium
(hyperstriatum – Wulst)
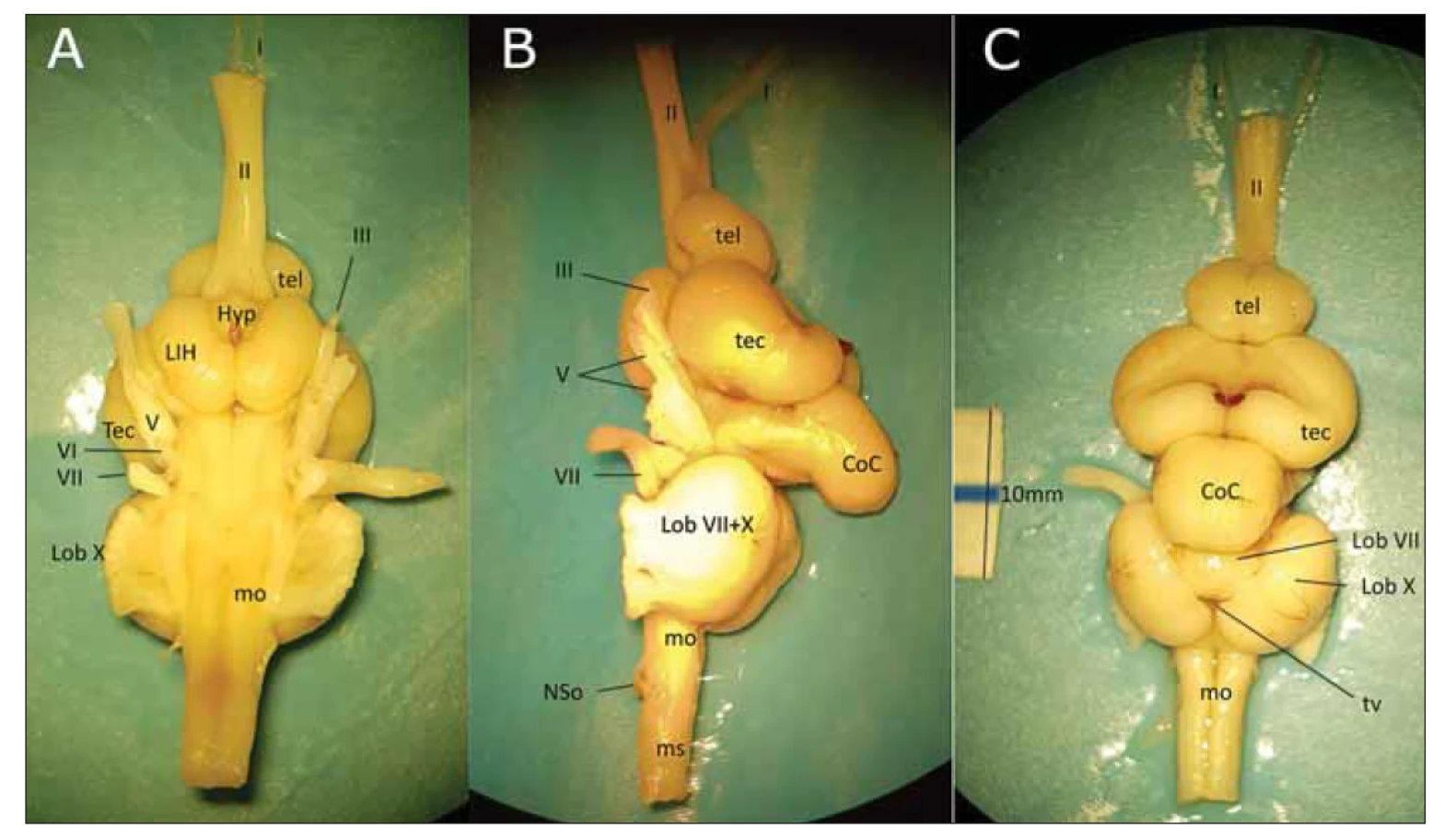
Dále jsme preparovali mozek kapra obecného (Cyprinus carpio) (obr. 8). U ryby v rhombencefalu prominují lobus facialis a lobus vagalis, velké tectum opticum překrývá diencefalon, telencefalon je velice malý a je uložen anteriorně nad vnořením čichového a zrakového traktu. U všech kaprovitých ryb se nachází výrazná párová vyvýšenina v kaudolaterální části spodiny IV. komory – lobus vagalis [19]. Je podmíněna chuťovými jádry nervus vagus (X) a nervus glossopharyngeus (IX). Uvnitř každého lobus vagalis je devět vrstev šedé hmoty složitě organizovaných v chuťové jádro. V jeho různých vrstvách končí senzitivní vlákna přivádějící ventrolaterální části X a IX chuťové informace z tlamy, z patrového orgánu a žaber. Nespecifická viscerocepce běží dorzálními částmi X a IX a končí v kaudální nepárové části ncl. tractus solitarii. Rostromediálně od lobi vagales se nachází lobus facialis, vyvýšenina daná chuťovým jádrem n. facialis (VII). Do něj přicházejí chuťová vlákna vedená ventrolaterální částí lícního nervu ze rtů, přední části patra, pektorální ploutve prozkoumávající dno rybníka i z celého povrchu těla. Tam všude má kapr chuťové pohárky. V tomto jádru se informace z rostrálních částí těla dostávají rostrálně a z kaudálních kaudálně. Vlákna z lobus vagalis míří do somatomotorických jader v oblongátě a zabezpečují různé krmivé a posturální reflexy. Vlákna z lobus facialis přivádějí informace do ncl. spinotrigeminalis funicularis, kde se u kapra integrují taktilní a chuťové informace. Ascendentní dráhy z obou lobi vagales míří do sekundárního chuťového jádra v pontu a odtud část z lobus vagalis do torus lateralis v mezencefalu (homolog savčího ncl. parabrachialis) a část z lobus facialis do lobus inferior hypothalami (savčí laterální hypotalamus). Tyto dvě struktury jsou vzájemně propojeny a projikují se do další úrovně mozku, do ncl. posterior thalami (část pregromerulárního komplexu pro ryby specifického tuberculum posterius), který zpracovanou informaci posílá do pallia. Terciální chuťové jádro umístěné v jiné části pregromerulárního komplexu (homolog savčího dorzálního talamu) získává informace ze sekundárního jádra a eferenty vysílá do lobus inferior hypothalami i do torus lateralis. Chuťový smysl je u kostnatých ryb, hlavně kaprovitých, skvěle vyvinut, což můžeme vidět na první pohled. Lobus vagalis a lobus facialis je velice vyklenutý. Pravděpodobně to vědí i rybáři chystající na tyto ryby návnadu. Informace z mechanoreceptorů postranní čáry jdou u kostnatých ryb do eminentia granularis mozečku a do laterálního sloupce jader na spodině IV. komory. Tento sloupec jader je dorzálně od jader VIII. hlavového nervu (oktaválních jader). Pokud mají ryby i elektroreceptory, končí nervy, které je zásobují, dorzálně od jader pro mechanoreceptory z postranní čáry. Z jader pokračuje tato senzitivní dráha oboustranně (hlavně kontralaterálně) pomocí lemniscus lateralis do torus semicircularis v tectum mesencephali a po přepojení do jader preglomerolárního komplexu pokračuje do pallia. Ryby nemají vyvinuté zevní a střední ucho, ale vnitřní blanitý labyrint reaguje jak na změnu polohy, tak na zvuk. Informace je vedena VIII. hlavovým nervem do dlouhé oktavální kolumny obsahující čtyři jádra a ležící ventrálně od jader pro informace z postranní čáry. Odtud dráha pokračuje laterálním lemniskem do torus semicircularis (homolog savčího colliculus inferior), potom do jader preglomerulárního komplexu a z nich do pallia. Motorická jádra na spodině IV. komory slouží k přijímání potravy a polykání, reakci na sluchové a zrakové podněty a k pohybům očí. U kapra přicházejí chuťové informace z lobus facialis do motorických jader n. trigeminus (V) a facialis (VII), k nim se přidávají vstupy ze senzitivního jádra V, z mozečku a tekta. To vše slouží ke koordinaci otvírání a zavírání tlamy při reakci na zevní podnět. Přidává se i informace z hypotalamu a retikulární formace. Polykání potravy je zabezpečeno činností nucleus ambiguus, společného motorického jádra pro IX, X a nervus accessorius, které je pod vlivem lobus vagalis. U suchozemských zvířat je třeba potravu před spolknutím zvlhčit, kvůli čemuž dochází k rozvoji jader inervujících slinné žlázy – ncl. salivatorius superior a inferior, poté promíchat se slinami a posunout do hltanu, tudíž se rozvíjí ncl. nervi hypoglossi.
I – tractus olfactorius; II – tractus opticus; III – n. oculomotorius; V – n. trigeminus; VI – n. abducens; VII – n. facialis; CoC – corpus cerebelli;
Hyp – hypothalamus; LIH – lobus inferior hypothalami; tec – tectum opticum; tel – telencephalon; Lob VII – lobus facialis; Lob X – lobus vagalis;
mo – medulla oblongata; ms – medulla spinalis; NSo – nervus spinooccipitalis; tv – tela ventriculi
Fig. 8. The brain of the carp – (A) ventral view; (B) lateral view; (C) posterior view, including the 10-mm measurement.
I – tractus olfactorius; II – tractus opticus; III – n. oculomotorius; V – n. trigeminus; VI – n. abducens; VII – n. facialis; CoC – corpus cerebelli;
Hyp – hypothalamus; LIH – lobus inferior hypothalami; tec – tectum opticum; tel – telencephalon; Lob VII – lobus facialis; Lob X – lobus vagalis;
mo – medulla oblongata; ms – medulla spinalis; NSo – nervus spinooccipitalis; tv – tela ventriculi
Vraťme se však zpět k motorickému systému. Motorika ryb je založena na kmenových jádrech a vliv telencefala se na kmen přenáší nepřímo [26]. Pokud však budeme hledat paralely a také rozdíly v uspořádání motorického systému, pomůže nám to lépe si zapamatovat funkční souvislosti v lidské neuroanatomii. Jako příklad uveďme, že u ryby jsou v předním míšním rohu motoneurony pro svaly ploutve na periferii a motoneurony pro svaly trupu jsou umístěny centrálně. U člověka je situace podobná, jen se „komplikuje“ tím, že pro svaly končetin (ploutví) jsou laterálně (na periferii), pro svaly trupu/krku mediálně, motoneurony pro flexory dorzálně a extensory ventrálně. Co se týče motorických drah, ryby mají vyvinutou vestibulospinální dráhu (tzv. oktavální). Oktavální je nazývána, protože hlavním aferentem vestibulárních jader je nervus vestibularis (osmý nerv). Rozlišujeme laterální vestibulární dráhu, která se nekříží a vychází z Deitersova jádra (laterálního), a oboustranně končící mediální dráhu, vycházející z mediálních jader. Deitersovo jádro také dostává aferentaci z Purkyňových buněk mozečku, jehož podsoučástí je ncl. medialis rhombencephalico-octavolaterální oblasti – primárního centra postranní čáry (mechanorecepční orgán ryb a larev obojživelníků). Podobně jako u savců bude sloužit motorické kontrole v závislosti na vnímání rovnováhy. Další dráhou ryb je retikulospinální dráha vycházející z jader od tegmenta mezencefala po prodlouženou míchu. Pozor, dostáváme se opět k naší predátorské teorii! Mezi retikulární centra u ryb, některých ocasatých obojživelníků a žab patří 2 gigantické Mauthnerovy buňky (pouze jedna na každé straně kmene), sloužící k bleskové únikové odpovědi, při které se ryba obloukovitě prohne hlavou směrem od podnětu (obr. 9). Na těle a laterálních dendritech těchto skoro 1 mm dlouhých buněk je několik set tisíc synapsí přivádějících podněty ze zrakové i sluchové dráhy, vestibulárního ústrojí, z postranní čáry, tekta a mozečku. Z Mauthnerovy buňky odstupuje jediný, tlustý a myelinizovaný axon, kříží střední čáru, sestupuje v laterálních míšních provazcích a aktivuje descendentní interneurony, motoneurony i komisurální buňky, které inhibují motoneurony na opačné straně. Reakce je blesková. Od akustického nebo taktilního stimulu po aktivaci Mauthnerových buněk uplyne 5–10 ms a po dalších 2 ms dojde ke svalové kontrakci. Škoda, že se takové buňky nezachovaly i u nás. U ryb byla také popsána role Mauthnerových buněk při lovu, kdy dojde ke stejnému rychlému obloukovitému prohnutí, ovšem hlavou směrem k předpokládané kořisti. Pro běžné plavání a orientaci těla vzhledem k proudu a gravitaci slouží v mezencefalu a oblongátě uložené menší shluky velkých Müllerových buněk, jejichž silně myelinizované axony se většinou kříží a probíhají v předních míšních provazcích. Co se týče motoriky ovládající přímo svaly trupu a ocasu ryby, zde se aktivuje jádro mediálního longitudinálního fasciklu odpovídající ncl. interstitialis Cajali, tedy intersticiospinální dráhy koordinující u savců pouze pohyby očních bulbů a svalů krku. U ryb však pochopitelně tato dráha zasahuje do celého rozsahu míchy. Vyvinutější motorika (což je tedy motorika ploutví, rybích končetin) je řízena z pars magnocellularis ncl. ruber (Goldsteinovo jádro) a inervuje již zkříženě svaly ploutví. Zde nacházíme jasnou paralelu s rubrospinální dráhou zapojenou do laterálního motorického systému u savců. Aby situace nebyla tak jednoduchá, některé druhy ryb (např. pstruh duhový) mají i nezkříženou rubrospinální dráhu vycházející z parvocelullárního Nieuwenhuysova a Pouwelsova ncl. ruber. K výraznějšímu vývoji rubrospinální dráhy však dochází až po výstupu obratlovců z vody na souš, hlavně u plazů. U ryb nebylo zjištěno přímé telencefalospinální spojení. Telencefalon má však vliv na motoriku též, a to nepřímo, po přepojení přes kmenová centra, např. elektrickou stimulací dorzálního telencefala u slunečnice velkoploutvé (Lepomis macrochirus) z čeledi okounkovitých byly vyvolány okamžité pohybové mechanizmy sloužící ke stavbě hnízda.
Zeleně – obrovské Mauthnerovy neurony (tělo 1 x 0,1 mm) na nichž jsou statisíce synapsí. Při
podráždění (postranní čára, vestibulární ústrojí, sluch, zrak) přicházejícím z jedné strany dojde
k integraci signálů na Mauthnerově buňce příslušné strany a bleskovému vedení vzduchu
do míchy. Silně myelinizovaný axon se kříží a běží v laterálním provazci. Excituje kontralaterální
motoneurony (červeně), descendentní interneurony (modře) a kommissurální interneurony
(prázdná černá kolečka). To vše vyvolá stah svalů kontralaterálně. Descendentní interneurony
aktivují motoneurony a komissurální interneurony tlumí motoneurony, descendentní interneurony
i kommissurální interneurony na opačné straně (na té odkud se něco blíží). Výsledkem je
bleskové otočení ryby od podnětu za 10–20 ms. Šipkami jsou značeny elektrické synapse, čárkou
chemické.
Fig. 9. The diagram of engagement of Mauthner cells in rapid escape reaction of the fish
(C-start) – (A) drawing of the escape reaction phases; (B) top view of the transparent head
and front of the fish body. According to [27], drawn by Monika Němcová.
Green – giant Mauthner cells (the body diameter 1 x 0.1 mm) – end point of a hundred thousand
synapses. After excitation (lateral line organ, vestibular apparatus, hearing, vision) incoming
from one side, signals in the ipsilateral Mauthner cell integrate and the signal is conducted
to the spinal cord. Heavily myelinated axon decussates and runs in the lateral funiculus. It
excites contralateral motoneurons (red), descendent interneurons (blue), and commissural interneurons
(empty black dots). This all leads to contralateral muscle contraction. Descendent
interneurons activate motoneurons and commissural interneurons inhibit motoneurons, as
well as descendent interneurons and also commissural interneurons on the other side (to something
that draws them near). The result is a lightning quick turnaround of the fi sh with 10–
20-ms latency from the stimulus. The arrows are the electric synapses, and chemical synapses
are dashed lines.
![Schéma zapojení Mauthnerových buněk při rychlém úniku ryb (C-startu) – (A) nákres
fází úniku; (B) pohled shora na průhlednou hlavu a přední část těla ryby. Podle [27],
nakreslila Monika Němcová.<br>
Zeleně – obrovské Mauthnerovy neurony (tělo 1 x 0,1 mm) na nichž jsou statisíce synapsí. Při
podráždění (postranní čára, vestibulární ústrojí, sluch, zrak) přicházejícím z jedné strany dojde
k integraci signálů na Mauthnerově buňce příslušné strany a bleskovému vedení vzduchu
do míchy. Silně myelinizovaný axon se kříží a běží v laterálním provazci. Excituje kontralaterální
motoneurony (červeně), descendentní interneurony (modře) a kommissurální interneurony
(prázdná černá kolečka). To vše vyvolá stah svalů kontralaterálně. Descendentní interneurony
aktivují motoneurony a komissurální interneurony tlumí motoneurony, descendentní interneurony
i kommissurální interneurony na opačné straně (na té odkud se něco blíží). Výsledkem je
bleskové otočení ryby od podnětu za 10–20 ms. Šipkami jsou značeny elektrické synapse, čárkou
chemické.<br>
Fig. 9. The diagram of engagement of Mauthner cells in rapid escape reaction of the fish
(C-start) – (A) drawing of the escape reaction phases; (B) top view of the transparent head
and front of the fish body. According to [27], drawn by Monika Němcová.<br>
Green – giant Mauthner cells (the body diameter 1 x 0.1 mm) – end point of a hundred thousand
synapses. After excitation (lateral line organ, vestibular apparatus, hearing, vision) incoming
from one side, signals in the ipsilateral Mauthner cell integrate and the signal is conducted
to the spinal cord. Heavily myelinated axon decussates and runs in the lateral funiculus. It
excites contralateral motoneurons (red), descendent interneurons (blue), and commissural interneurons
(empty black dots). This all leads to contralateral muscle contraction. Descendent
interneurons activate motoneurons and commissural interneurons inhibit motoneurons, as
well as descendent interneurons and also commissural interneurons on the other side (to something
that draws them near). The result is a lightning quick turnaround of the fi sh with 10–
20-ms latency from the stimulus. The arrows are the electric synapses, and chemical synapses
are dashed lines.](https://www.prelekara.sk/media/cache/resolve/media_object_image_large/media/image_pdf/86dc0986075176c6be8ff492cebb3ef7.png)
Potud naše malá exkurze do tajů mozku kapra: kmen: Chordata, podkmen: Vertebrata, nadtřída: Osteichthyes, třída: Actinopterygii, řád: Cypriniformes, čeleď: Cyprinidae, rod: Cyprinus; kachny: kmen: Chordata, podkmen: Vertebrata, nadtřída: Amniota, třída: Sauropsida, podtřída: Aves, řád: Anseriformes, čeleď: Anatidae, rod: Anas a králíka: kmen: Chordata, podkmen: Vertebrata, nadtřída: Amniota, třída: Synapsida, podtřída: Mammalia, řád: Lagomorpha, čeleď: Leporida, rod: Oryctolagus.
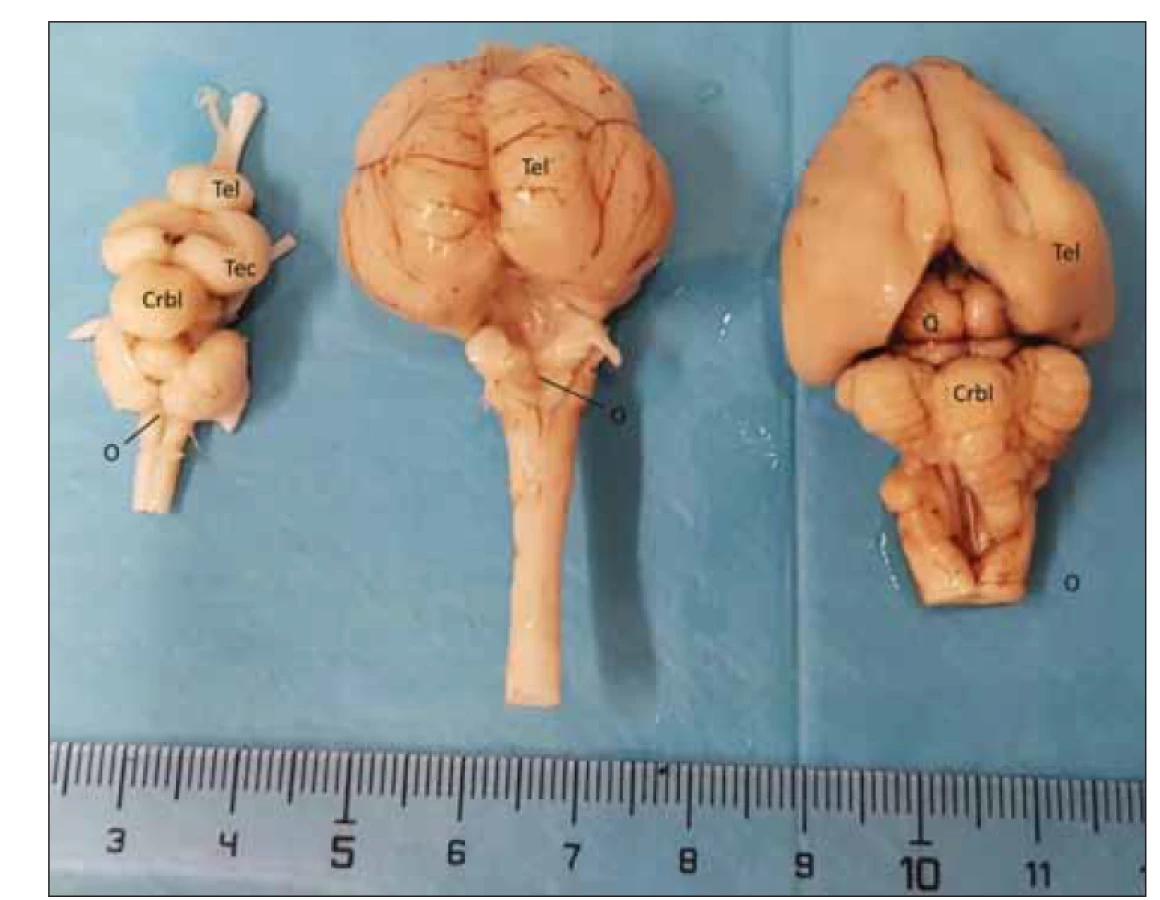
Na obr. 10 jsou porovnány mozky kapra, kachny a králíka z dorzální strany. Nápadné jsou malý telencefalon u ryby, vyklenutý Wulst na ptačím mozku a lissencefalní hemisféra králíka. Již Aristoteles (384–321 př. n. l.) se domníval, že zvíře má duši, a nabádal k pozorování krás přírody. V tomto smyslu apeloval na lidstvo také lékař a humanista Albert Schweitzer (1875–1965): „Snažme se udělat konec primitivní krutosti, s jakou zacházíme se zvířaty. Zvířata cítí bolest stejně jako my.“ Všechny naše preparáty byly získány ze zvířecích kadaverů ze zvířat zabitých pro lidskou obživu rukou farmáře nebo rybáře. Studium anatomie stavby mozku zvířat nám může pomoci pochopit složitost lidského mozku i důvody křížení mozkových drah, a proto jsme dané disekce prováděli. Během tohoto studia jsme se ke zvířecímu mozku chovali se stejnou úctou a pokorou jako to činíme při práci s mozkem lidským. Náš referát bychom rádi zakončili citací George Orwella z knihy Farma zvířat (1945): „Všechna zvířata jsou si rovna, ale některá jsou si rovnější“. Tato věta se bohužel týká jen nás lidí.
Crbl – cerebellum; O – obex; Q – lamina quadrigemina; Tec – tectum opticum; Tel – telecephalon
Fig. 10. Comparison of the brain of the carp (left), duck (middle), and rabbit (right), including
the 10-mm measurement.
Crbl – cerebellum; O – obex; Q – lamina quadrigemina; Tec – tectum opticum; Tel –
telencephalon
Grantová podpora
Podpořeno vnitřním grantem IG7 – 217111029. Tato práce byla částečně podpořena vnitřním grantem KZ. a. s. (IGA-KZ-2020-1-20).
Konflikt zájmů
Autoři deklarují, že v souvislosti s předmětem práce nemají žádný konflikt zájmů.
MUDr. Veronika Němcová, CSc.

MUDr. Veronika Němcová, CSc. se narodila 13. 6. 1963 v porodnici na pražské Štvanici. V roce 1981 odmaturovala na Gymnáziu Nad Štolou. Po roce v „nultém ročníku“ na ORL ve Všeobecné fakultní nemocnici na Praze 2 byla přijata na Fakultu všeobecného lékařství (dnešní 1. LF UK), kde v roce 1988 promovala. Nastoupila na Anatomický ústav 1. LF UK, kde pracovala již za studií, a zároveň sbírala zkušenosti na neurologické klinice. Po 2 letech klinickou praxi opustila a zapojila se do výzkumu retikulární formace mozkového kmene v laboratoři prof. Petrovického. Absolvovala 2 dvouměsíční pobyty v neurolaboratoři na Katholieke Universiteit v Nijmegenu u prof. ten Donkelaara. V roce 1997 obhájila práci Nucleus laterodorsalis tegmenti a další struktury pontomesencephalického přechodu a získala titul CSc. Zajímá ji cytoarchitektonika a spoje jader retikulární formace se zaměřením na NADPHd pozitivní neurony. Od roku 2009 se zabývala volumometrií hippokampu a amygdaly u narkolepsie ve spolupráci s Radiologickou a Neurologickou klinikou 1. LF UK. Spolupracuje i s neurochirurgickými pracovišti v ÚVN a Masarykově nemocnici v Ústí nad Labem v oblasti studia periferních nervů a při preparacích mozkových spojů a cév. Na Web of Science má 7 původních článků, 25 citací a H-index 2, v ORCID 15 prací. Podílí se na tvorbě a překladech učebnic, prezentací a výukových videí. Jako odborná asistentka přednáší anatomii, vede výuku pitevních cvičení, praktik a je garantem volitelného předmětu Dráhy CNS na 1. LF UK. Má tři už dospělé děti, ráda chodí do přírody a plave.
Redakční rada potvrzuje, že rukopis práce splnil ICMJE kritéria pro publikace zasílané do biomedicínských časopisů.
The Editorial Board declares that the manuscript met the ICMJE “uniform requirements” for biomedical papers.
MUDr. Veronika Němcová, CSc.
Anatomický ústav
1. LF UK
U Nemocnice 3
128 00 Praha
e-mail: veronika.nemcova@lf1.cuni.cz
Přijato k recenzi: 24. 2. 2021
Přijato do tisku: 22. 7. 2021
Sources
1. Ramón y Cajal S. Texture of the nervous system of man and the vertebrates. J Neurol Neurosurg Psychiatry 2001; 70 (3): 421. doi: 10.1136/jnnp.70.3.421c.
2. Mora C, Velásquez C, Martino J. The neural pathway midline crossing theory: a historical analysis of Santiago Rámon y Cajal’s contribution on cerebral localization and on contralateral forebrain organization. Neurosurg Focus 2019; 47 (3): E10. doi: 10.3171/2019.6. FOCUS19341.
3. Vulliemoz S, Raineteau O, Jabaudon D. Reaching beyond the midline: why are human brains cross wired? Lancet Neurol 2005; 4 (2): 87–99. doi: 10.1016/S1474-4422 (05) 00990-7.
4. Cajal RM. Estructura del kiasma óptico y teoría general de los entrecruzamientos de las vías nerviosas. Rev Trim Micrografica 1898; 3 : 15–66.
5. Kinsbourne M. Somatic twist: A model for the evolution of decussation. Neuropsychology 2013; 27 (5): 511–515. doi: 10.1037/a0033662.
6. de Lussanet MHE, Osse JWM. An ancestral axial twist explains the contralateral forebrain and the optic chiasm in vertebrates. Animal Biol 2012; 62 (2): 193–216. doi: 10.1163/157075611X617102.
7. de Lussanet MH, Osse JW. Decussation as an axial twist: a comment on Kinsbourne (2013). Neuropsychology 2015; 29 (5): 713–714. doi: 10.1037/neu0000163.
8. Zalocusky K. Ask a neuroscientist: why does the nervous system decussate? [online]. Available from URL: https: //neuroscience.stanford.edu/news/ask-neuroscientist-why-does-nervous-system-decussate.
9. Shinbrot T, Young W. Why decussate? Topological constraints on 3D wiring. Anat Rec (Hoboken) 2008; 291 (10): 1278–1292. doi: 10.1002/ar.20731.
10. Long H, Sabatier C, Ma L et al. Conserved roles for slit and Robo proteins in midline commissural axon guidance. Neuron 2004; 42 (2): 213–223. doi: 10.1016/s0896-6273 (04) 00179-5.
11. Sabatier C, Plump AS, Ma L et al. The divergent Robo family protein rig-1/Robo3 is a negative regulator of slit responsiveness required for midline crossing by commissural axons. Cell 2004; 117 (2): 157–169. doi: 10.1016/s0092-8674 (04) 00303-4.
12. Neuhaus-Follini A, Bashaw GJ. Crossing the embryonic midline: molecular mechanisms regulating axon responsiveness at an intermediate target. Wiley Interdiscip Rev Dev Biol 2015; 4 (4): 377–389. doi: 10.1002/wdev.185.
13. Gunderson CH, Solitare GB. Mirror movements in patients with the Klippel-Feil syndrome. Neuropathologic observations. Arch Neurol 1968; 18 (6): 675–679. doi: 10.1001/archneur.1968.00470360097009.
14. Danek A, Heye B, Schroedter R. Cortically evoked motor responses in patients with Xp22.3-linked Kallmann’s syndrome and in female gene carriers. Ann Neurol 1992; 31 (3): 299–304. doi: 10.1002/ana.410310312.
15. Mayston MJ, Harrison LM, Quinton R et al. Mirror movements in X-linked Kallmann’s syndrome. I. A neurophysiological study. Brain 1997; 120 (Pt 7): 1199–1216. doi: 10.1093/brain/120.7.1199.
16. Farmer SF, Harrison LM, Mayston MJ et al. Abnormal cortex-muscle interactions in subjects with X-linked Kallmann’s syndrome and mirror movements. Brain 2004; 127 (Pt 2): 385–397. doi: 10.1093/brain/awh047.
17. Jen JC, Chan WM, Bosley TM et al. Mutations in a human ROBO gene disrupt hindbrain axon pathway crossing and morphogenesis. Science 2004; 304 (5676): 1509–1513. doi: 10.1126/science.1096437.
18. Sharpe JA, Silversides JL, Blair RD. Familial paralysis of horizontal gaze: associated with pendular nystagmus, progressive scoliosis, and facial contraction with myokymia. Neurology 1975; 25 (11): 1035–1040. doi: 10.1212/wnl.25.11.1035.
19. Marillat V, Sabatier C, Failli V. The Slit receptor Rig-1/Robo3 controls midline crossing by hindbrain precerebellar neurons and axons. Neuron 2004; 43 (1): 69–79. doi: 10.1016/j.neuron.2004.06.018.
20. Sarnat HB, Netsky MG. Evolution of the nervous system. New York, NY: Oxford University Press 1974.
21. Butler A. Comparative vertebrate neuroanatomy evolution and adaptation. 2nd ed. Hoboken (NJ): Wiley-Interscience 2005.
22. Edinger L. Vorlesungen űber den Bau der Nervösen Zentralorgane des Menschen und der Tiere: fűr Ärzte und Studierende. Leipzig: Vogel 1908.
23. Kappers A, Huber CU, Crosby CG. The comparative anatomy of the nervous system of vertebrates including man. New York: Macmillan 1936.
24. Reiner A, Perkel DJ, Bruce LL. Revised nomenclature for avian telencephalon and some related brainstem nuclei. J Comp Neurol 2004; 473 (3): 377–414. doi: 10.1002/cne.20118.
25. Jarvis ED, Yu J, Rivas MV et al. Global view of the functional molecular organization of the avian cerebrum: Mirror images and functional columns. J Comp Neurol 2013; 521 (16): 3614–3665. doi: 10.1002/cne.23 404.
26. Yamamoto N, Nakayama T, Hagio H. Descending pathways to the spinal cord in teleosts incomparison with mammals, with special attention to rubrospinal pathways. Dev Growth Differ 2017; 59 (4): 188–193. doi: 10.1111/dgd.12355.
27. Fetcho JR. Spinal network of the mauthner cell (part 1 of 2). Brain Behav Evol 1991; 37 (5): 298–306. doi: 10.1159/000114367
Labels
Paediatric neurology Neurosurgery NeurologyArticle was published in
Czech and Slovak Neurology and Neurosurgery

2021 Issue 4
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Advances in the Treatment of Myasthenia Gravis on the Horizon
Most read in this issue
- COVID-19 related olfactory impairment – diagnostics, significance and treatment
- CANVAS – a newly identified genetic cause of late-onset ataxia. Description of the first cases in the Czech Republic
- Why do the nerve tracts decussate? Basic principles of the vertebrate brain organization
- COVID-19 associated myelitis – a case report of rare complication of severe SARS-CoV-2 infection