
Evoluce a evoluční teorie pro lékaře
X. Sebeuvědomování
Evolution and evolutionary theory for physicians
X.: Self-awareness.
Evolution of self-awareness is hypothetically based on
– ecological,
– interpersonal,
– extended,
– private, and
– conceptual self-knowledge.
Neuronal correlate of self-awareness might be brain´s default (intrinsic) network. Its core regions are
– ventromedial prefrontal cortex,
– posterior cingular/retrosplenial cortex,
– inferior parietal lobule,
– lateral temporal cortex,
– dorsal medial prefrontal cortex, and
– hippocampal formation.
High heritability exists for default-mode functional connectivity but its hubs are under dynamic cultural influences too.
Key words:
self-awareness, evolution, default network.
Authors:
F. Koukolík
Authors‘ workplace:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
Published in:
Prakt. Lék. 2010; 90(10): 571-576
Category:
Editorial
Overview
Evoluce sebeuvědomování je hypoteticky založena na vývoji
– ekologického,
– interpersonálního,
– rozšířeného,
– soukromého, a
– symbolického jáství.
Možným neuronálním korelátem sebeuvědomování je činnost mozkové implicitní (defaultní) sítě tvořené
– kůrou ventromediální prefrontální,
– zadní cingulární / retrosplenickou kůrou,
– kůrou lobulus parietalis inferior,
– kůrou zevní spánkovou,
– kůrou dorzální prefrontální, a
– hipokampem.
Heritabilita funkční konektivity intrinsické sítě je vysoká, avšak na její činnost působí i kulturní vlivy.
Klíčová slova:
sebeuvědomování, evoluce, intrinsická (default) síť lidského mozku.
Úvod
Pojem sebeuvědomování má několik významů:
1. Sebeuvědomování jako sebepoznávání.
Děti začnou poznávat samy sebe v zrcadle ve věku kolem 18 měsíců, přibližně ve stejné době začnou užívat osobní zájmena. Plně vyvinuté jáství lze pravděpodobně počítat od věku 36–48 měsíců, od doby, v níž je plně vyvinutá mentalizace, to je schopnost rozlišit, že i druhý jedinec má niterné stavy, například o něčem přemýšlí, něco prožívá, má nějaký záměr (12). V zrcadle se poznávají šimpanzi, delfíni, sloni i straky (19).
2. Sebeuvědomování jako vědomí o vědomí,
například vědomí o zrakovém, sluchovém nebo somatosenzorickém vědomí, rozlišování „já“ od „někdo jiný“ v průběhu mentalizace.
3. Sebeuvědomování jako znalost sama sebe v sociálním slova smyslu,
vědomí sama sebe jako člena lidských skupin. V tomto smyslu se sebeuvědomování vyvíjí a diferencuje celý život.
Vývoj sebeuvědomování
Hypotetický vývoj lidského sebeuvědomování („jáství“) v tradičním vývojovém modelu našeho druhu je v tab. 1. Hypotéza (14) předpokládá, že jáství má pět domén:
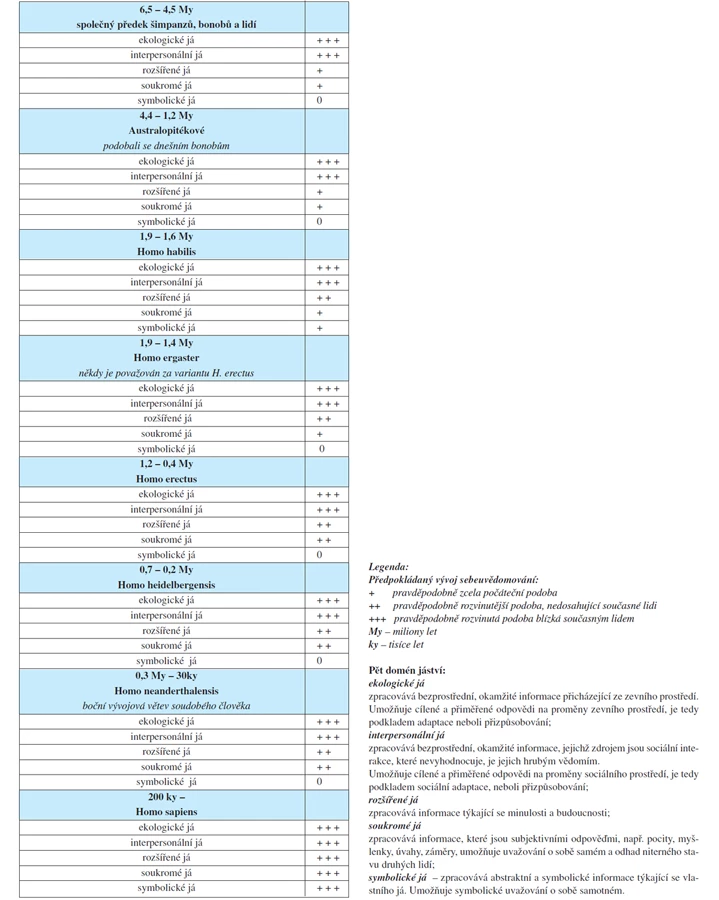
- ekologické já
zpracovává bezprostřední, okamžité informace přicházející ze zevního prostředí. Umožňuje cílené a přiměřené odpovědi na proměny zevního prostředí, je tedy podkladem adaptace, neboli přizpůsobování;
- interpersonální já
zpracovává bezprostřední, okamžité informace, jejichž zdrojem jsou sociální interakce, které nevyhodnocuje, je jejich hrubým vědomím. Umožňuje cílené a přiměřené odpovědi na proměny sociálního prostředí, je tedy podkladem sociální adaptace, neboli přizpůsobování;
- rozšířené já
zpracovává informace týkající se minulosti a budoucnosti;
- soukromé já
zpracovává informace, které jsou subjektivními odpověďmi, např. pocity, myšlenky, úvahy, záměry, umožňuje uvažování o sobě samém a odhad niterného stavu druhých lidí;
- symbolické
zpracovává abstraktní a symbolické informace týkající se vlastního já. Umožňuje symbolické uvažování o sobě samotném.
Implicitní síť
Neuronální korelát sebeuvědomování začal být odkrýván objevem implicitní (default) sítě lidského mozku (20). Teoretický předpoklad říkal, že činnost lidského mozku ponechaného v „klidu“, to znamená jedinec v bdělém stavu, tělo adaptované na tlak podložky, zavřené oči, žádné nové zvuky, kromě „šumu pozadí“, bude nepředvídatelně kolísat, úměrně tomu, co si budou lidé představovat, nebo na co budou myslet, jak budou sami se sebou ve vlastním nitru mluvit.
Zjištění, že mozek má, měřeno extrakční frakcí kyslíku, „bazální stav“ (baseline state), bylo překvapující. Extrakční frakce kyslíku se určuje poměrem kyslíku spotřebovaného mozkem ku kyslíku, který dodal krevní oběh v bdělém a klidovém stavu, což lze zjistit pozitronovou emisní tomografií a funkční magnetickou rezonancí. Jakékoli změny činnosti mozku lze určit jako odchylky od tohoto bazálního stavu.
Jakmile se mozek zatíží nějakou úlohou, poklesne takto měřená neuronální aktivita v systému oblastí, pro který bylo vytvořeno označení implicitní (default) síť (intrinsic, default network).
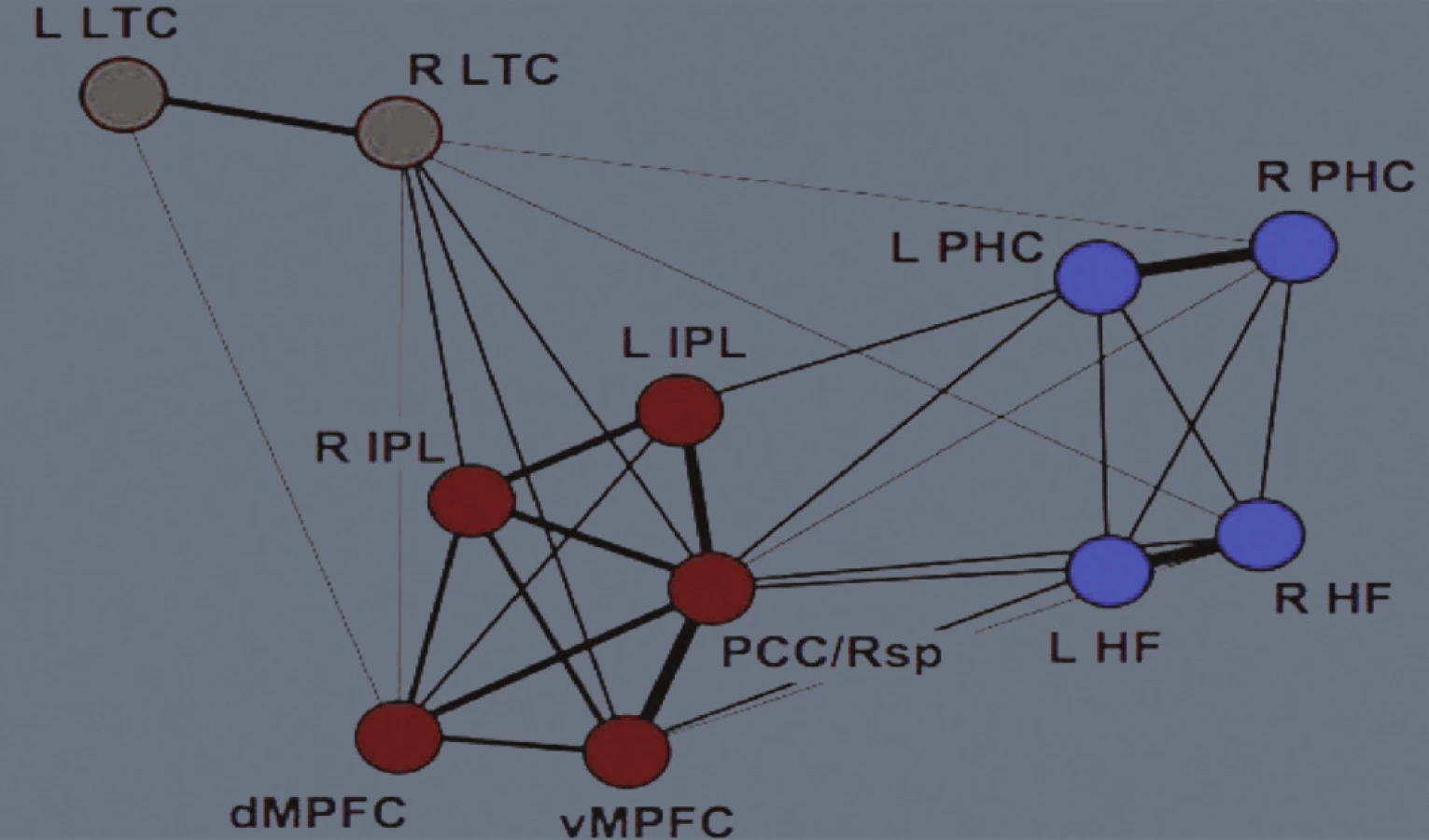
Klíčovými uzly „těžišti“, („nábami“, angl. hubs) implicitní sítě jsou
- ventromediální prefrontální kůra (Brodmanova area, BA, 24, 10, 32,
- zadní cingulární a retrosplenická kůra (BA 29/30, 23/31),
- lobulus parietalis inferior (BA 39, 40),
- laterální spánková kůra (BA 21),
- dorsomediální prefrontální kůra (BA 24, 32, 9, zadní část B 10, a
- hipokampální formace včetně vlastního hipokampu (obr. 1 a 2) ( 2).
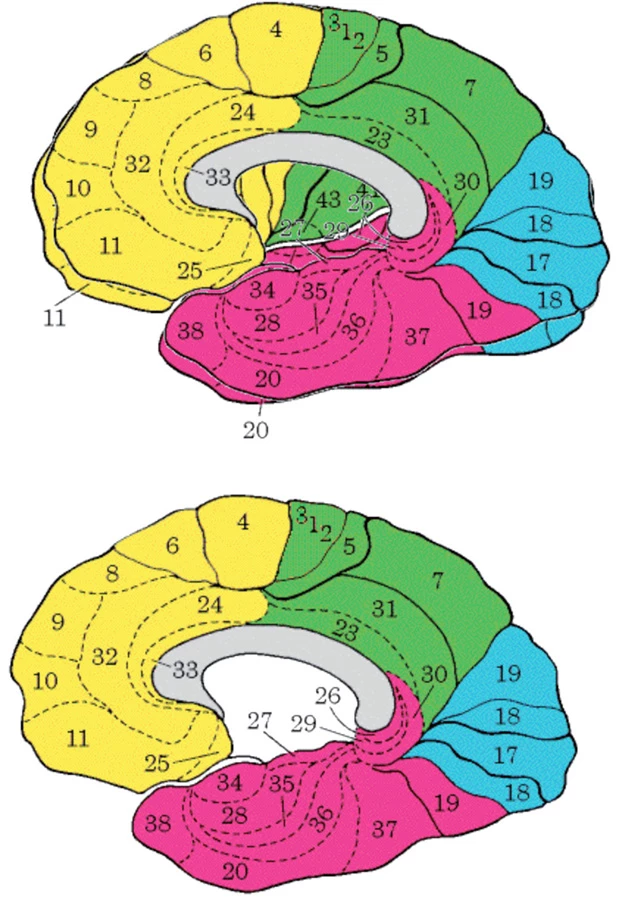
Aktivace této sítě je tedy opačná, antiparalelní, než je aktivace sítí zatížených například percepčními, v experimentech obvykle zrakovými, případně kognitivními úlohami.
Zjednodušeně řečeno: jakmile musí mozek zpracovávat informace přicházející ze zevního nebo vnitřního prostředí, přestane se věnovat sám sobě.
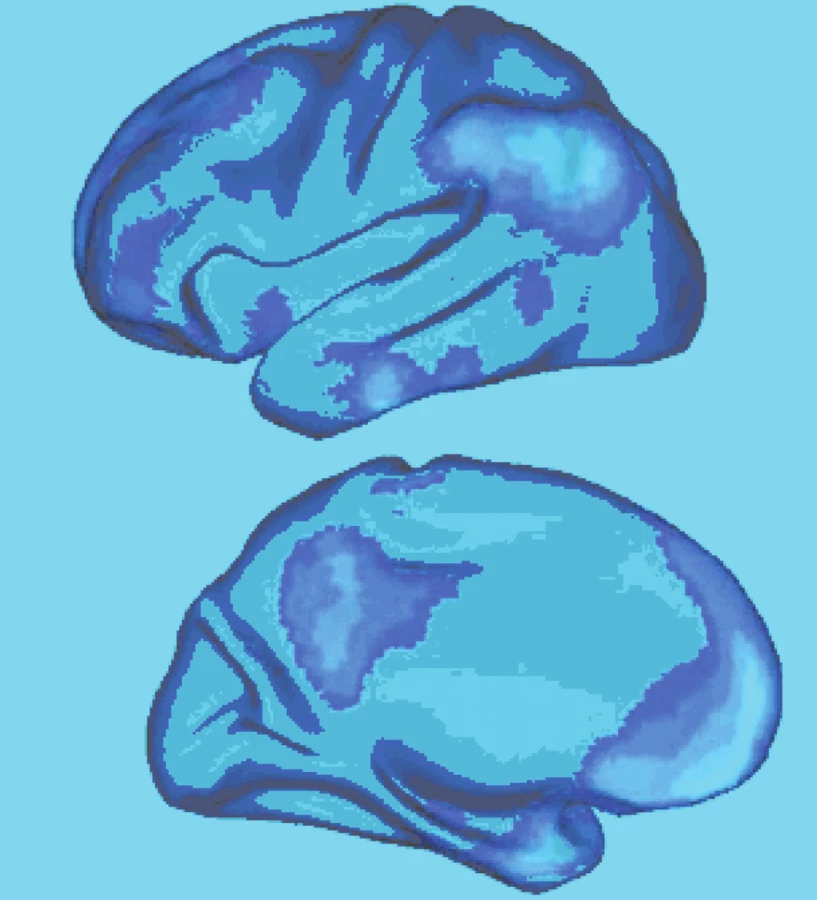
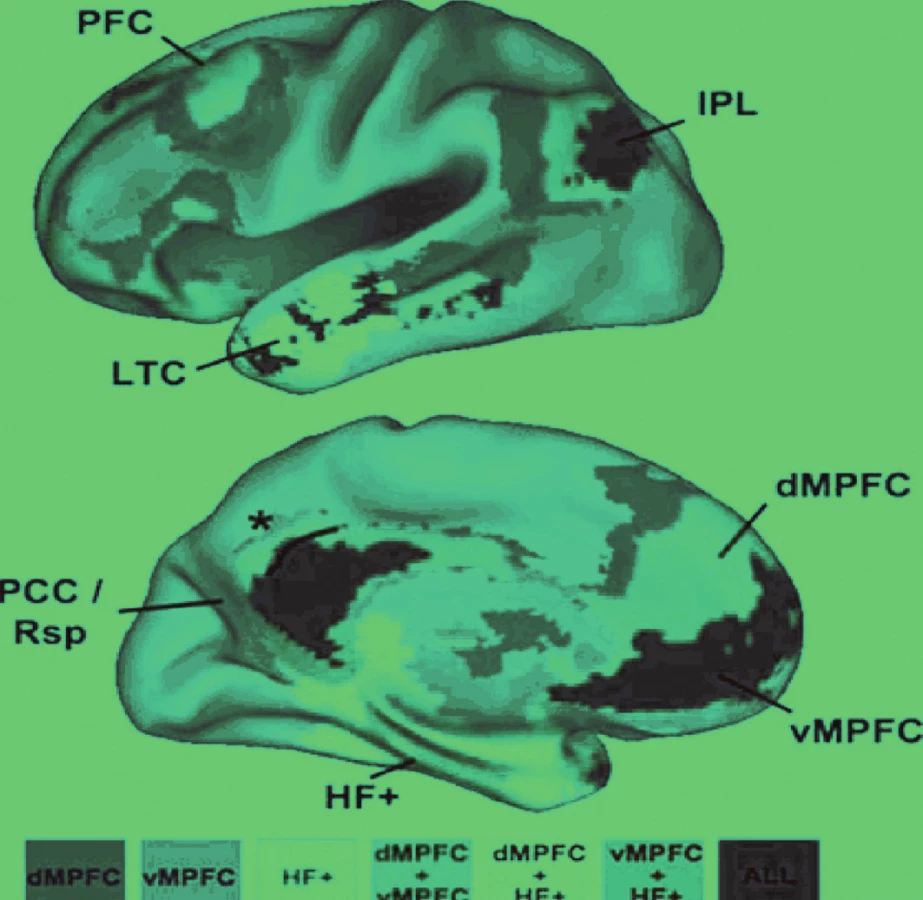
Jednotlivé uzly sítě aktivují společně v průběhu „pasivních“ stavů, onoho „klidu“, jejich činnost je korelovaná a jsou vzájemně přímo a nepřímo propojené. Síť samotnou je možné rozdělit do tří funkčních oddílů (obr. 3). Mohutnost propojení, funkční korelace, jednotlivých oddílů sítě je různá (obr. 4).
Jestliže jsme ponecháni sami sobě a nezvládáme problém vyžadující plnou pozornost, naše vědomí („mysl“, duše“, „já“…) putuje z jednoho obsahu k jinému: „spatřujeme“ minulé události, předvídáme budoucí, přemítáme o událostech a světech, které jsou mimo naše bezprostřední okolí. William James (1890) (10) to nazval proudem vědomí.
V současné době se mluví o spontánní kognici, putování mysli, myšlenkách nezávislých na podnětech (stimulus independent thoughts), což je jev popsaný antickými filozofy a z pozorování sebe sama známý pravděpodobně každému člověku, kromě jiného v podobě nazvané denní snění. Až 96 % lidí říká, že má tuto zkušenost každý den.
Experiment dokládá, že myšlenky nezávislé na podnětech odpovídají aktivitě implicitní sítě, zvláště mediální prefrontální a zadní cingulární kůry společně s kůrou retrosplenickou (obr. 3) (16). Aktivita implicitní sítě také odpovídá okamžitým výpadům pozornosti, chvílím, kdy naše vědomí „odputuje“ z právě řešené úlohy (15), záleží však na experimentálním zadání, protože se někdy může naopak zvýšit (7).
Aktivita intrinsické sítě kolísá, spontánně poměrně pomalu roste a klesá. Příkladem je korelovaný růst aktivity mediální prefrontální a zadní cingulární a spolu s ní restrosplenické kůry v pětiminutových intervalech při pozorování zaměřovacího kříže. Obě korové oblasti jsou vzdálené a v odlišných cévních povodích, takže korelace jejich aktivity je neuronální, nejde o měření šumu (4). Jakmile začnou lidé řešit nějakou kognitivní úlohu a jejich mysl „duše“, „jáství, přestane „putovat“, spontánní aktivita implicitní sítě klesá.
Z experimentů plyne, že funkcí intrinsické sítě může být
- niterný mentální život značně odloučený od zevního světa, tedy sebeprožívání, „niterné jáství“, „niterný aspekt duše“,
- podpora sledování zevního prostředí, jestliže je utlumena zaměřená, neboli fokalizovaná pozornost (obr. 5).
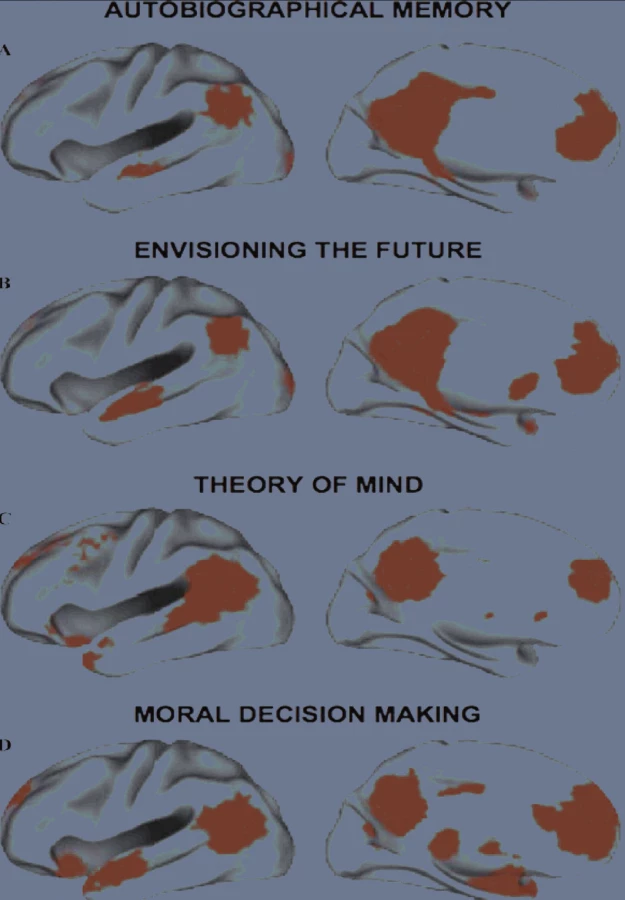
Implicitní síť totiž aktivují značně rozmanité úlohy:
- zatížení autobiografické paměti, jakmile si vybavujeme osobní události z minulosti,
- předvídání budoucnosti (tomu se někdy říká paměť pro budoucí události),
- mentalizace, neboli „čtení“ niterných duševních stavů druhého člověka, ale i morální rozhodování (1, 21, 6).
Implicitní síť, které se tedy přičítá autoreferenční, introspektivní aktivita (synonyma jsou niterné vyprávění, neboli internal narrative, autobiografické já, sebe-projekce čili self-projection) vyzrává v průběhu dospívání. U dětí ve věku 7–9 let jsou její těžiště propojena jen velmi slabě (3).
Supekarova skupina (23, 24) vyšetřovala vyzrávání implicitní sítě třemi metodami. Dokázala vývojové změny strukturálního i funkčního zapojení, které nejsou ve všech uzlech sítě uniformní. V souhlasu s předchozím popisem skupina doložila, že nejslabší je propojení mediální prefrontální kůry a zadní cingulární kůry. Kromě toho u dětí neexistuje nebo je velmi slabé strukturální propojení zadní cingulární kůry a levé mediální temporální kůry, přestože funkční konektivita je stejná jako u dospělých lidí. Z toho plyne, že funkční konektivita předchází konektivitu strukturální. Je zřejmé, že právě tato konektivita je jedním z podkladů vyzrávání autoreferenční sociální kognice v průběhu dospívání.
Činnost mediálního frontálního a cingulárního těžiště intrinsické sítě je možné disociovat. Pokusné osoby byly požádány, aby myslely na své naděje a aspirace, nebo na své povinnost a závazky. Činnost mozku při těchto sebereflexích lze porovnat s činností mozku při zátěži, která sebereflexivní není.
Sebereflexivní úlohy v porovnání se zátěží, která sebereflexivní není, podle očekávání zvyšují činnost ventromediální prefrontální a zadní cingulární kůry (včetně kůry precuneu). Jestliže pokusné osoby uvažovaly o nadějích a aspiracích, zvýšila se činnost mediální prefrontální a přední cingulární kůry. Jestliže uvažovaly o povinnostech a závazcích, vzrostla činnost zadní cingulární kůry a kůry precuneu (11).
Činnost implicitní sítě se je odlišná u autistů, při schizofrenii i Alzheimerově nemoci (2). Zajímavá je odchylná činnost sítě při velké depresi: jestliže se tito pacienti začnou dívat na obrázky s negativním emočním obsahem a vyhodnocovat je, pak narozdíl od lidí zdravých řada oblastí jejich implicitní sítě činnost neutlumí, v dalších oblastech aktivita sítě dokonce stoupne (22).
Vanhaudenhyuse et al. (25) testovala konektivitu implicitní sítě u nekomunikujících pacientů s poškozením mozku. Korelace konektivity všech oblastí sítě a klinického stavu vědomí při porovnání lidí zdravých, lidí se syndromem uzamčení (locked-in syndrome), lidí s minimálním vědomím, lidi ve vegetativním stavu a lidí v komatu, byla negativní.
U lidí ve stavu minimálního vědomí (to je varianta permanentního vegetativního stavu s částečně zachovanými některými korovými funkcemi (13), byla významně vyšší konektivita precuneu.
Konektivita pacientů se syndromem uzamčení se od konektivity zdravých lidí nelišila.
Konektivita intrinsické sítě se podílí na empatii. Otti et al. (17) předváděli zdravým dobrovolníkům obrázky lidských končetin v bolestivé a nebolestivé situaci. Přitom porovnali činnost intrinsické sítě jejich mozku v klidovém stavu a při těchto dvou druzích zátěže. Po vyšetření ve skeneru pokusné osoby vyhodnocovaly intenzitu pozorované bolesti tak, jako by ji prožily samy. Vyšší míře tohoto hodnocení odpovídala vyšší míra aktivity levostranné mediální prefrontální kůry (Brodmanova area 32).
Souhrnně řečeno: implicitní síť je soubor podsystémů v interakci. Podsystém, jehož uzlem je vnitřní spánková kůra, zajišťuje informace z předchozích zkušeností uložených jako paměťové záznamy. Mediální prefrontální podsystém umožňuje jejich pružné adaptivní užívání. Oba tyto podsystémy konvergují v integračním uzlu tvořeném zadní cingulární kůrou. Implicitní síť umožňuje adaptivní užití minulé zkušenosti při plánování budoucnosti (2).
Geny i kulturní vlivy
Implicitní síť byla doložena u opic v anestézii (26). Lze tedy předpokládat její existenci u společného předka opic a člověka. Průkaz genetického ovlivnění stavby a činnosti intrinsické sítě znamená vhled do jejích evolučních kořenů. Glahn et al. (5) v tomto směru vyšetřil 333 jedinců z 29 náhodně volených rodokmenů. Dědivost (heritabilita; P1) funkční konektivity intrinsické sítě je vysoká a byla zjištěna ve výši 0, 426 (± 0,17; p = 0,0046). Geny rovněž ovlivňují denzitu šedé hmoty sítě, takže ovlivňují jak její stavbu tak její funkci.
Konektivitu implicitní sítě ovlivňují kulturní vlivy. Individualisté chápou sami sebe jako autonomní stabilní bytosti oddělené od druhých lidí a od prostředí, kolektivisté chápou sami sebe jako dynamické jednotky trvale určované svými sociální vztahy a souvislostmi. Individualistické a kolektivistické pojetí sama sebe lze určit Singelisovou stupnicí. Experimenty ověřily, že individualistické a kolektivistické pojetí sama sebe odlišuje aktivita přední části mediální prefrontální a zadní cingulární kůry (8, 9).
Kromě jiného z nich plyne, že oddělování evolučních, tedy genetických, neuronálních a kulturních vlivů na lidské chování je možné jen didakticky.
Jaderné jáství jako evoluční jev
Není nutné zdůrazňovat, že povaha „jáství“, sebeuvědomování byla jedním z ústředních problémů filozofie. Staré otázky se ptají, zda mají sebeuvědomování i zvířata, a jestliže je mají, v jaké míře je s nimi lidé sdílejí, podobně jako sdílíme třeba procesy, které jsou podkladem vidění nebo motoriky.
Pankseppova skupina dokazuje v řadě studií (přehled 2009) existenci „jaderného jáství“ (core self), které mají lidé společné s živočichy. Jaderné jáství je popisováno jako specifický způsob interakce organismu s prostředím, jenž je projevem činnosti vymezených neuronálních sítí.
Jaderné jáství je hierarchicky uspořádaný systém. Jeho nejnižší úroveň odpovídá interocepci. V každé „krychličce“ lidských tkání, jejíž hrana měří 15 mikrometrů, jsou zakončení C a A-delta vláken, která rozlišují
- teplotu,
- mechanický stres,
- výši pH,
- hypoxii,
- hyperkapnii,
- hypoglykemii,
- prasklé buňky,
- parazity pronikající kůží,
- aktivaci žírných buněk,
- imunitní aktivitu,
- hormonální aktivitu, a
- senzuální dotyk.
Mozek tímto způsobem trvale mapuje vnitřní prostředí, jeho mapy lze chápat jako „vegetativní jáství“. Mapování probíhá v kmenových strukturách, mezimozku a insulách. Viscerální a somatosenzorické informace tvoří primitivní jáství („proto-self“), které můžeme mít společné s obratlovci.
Vyšší úroveň zpracovávání probíhá v subkortikální síti střední čáry tvořené šedí kolem mokovodu, horními hrbolky čtverohrbolí, sousedícími motorickými oblastmi mesencefala, area preoptica, hypothalamem a thalamem. Tvoří implicitní a afektivní podoby jaderného jáství. Tuto úroveň můžeme mít společnou se savci.
Nejvyšší úrovní je zpracovávání informací v nekortikálních oblastech tvořících explicitní a kognitivní podoby jáství, úroveň, jejíž složky máme společné s se sociálně žijícími primáty.
Poselství této myšlenkové školy v rozporu s klasickou racionalistickou tradicí říká, že „jáství“ je evoluční kontinuum, jehož povaha je z valné části „vegetativní“, somatosenzorická, afektivní a nevědomá.
Ontogeneze sebeuvědomování
Ontogeneze sebeuvědomování začíná s vývojem systému
- mentalizace,
- empatie,
- jazyka, a
- řídících funkcí.
Ve věku 0–9 měsíců se u dětí nejprve vyvíjejí tři „detektory“:
- detektor záměru,
- detektor směru pohledu, a
- detektor emocí,
což jsou funkční systémy umožňující nejmenším dětem určit co se děje v osobě a s osobou, která o ně systematicky pečuje. V rozmezí 9–14 měsíců se na základě vývoje „detektorů“ objevuje mechanismus sdílené pozornosti. Kolem 14. měsíce se na základě předchozích vývojových stupňů objevuje schopnost empatizovat, kolem 48. měsíce je plně vyvinuta mentalizace (theory of mind; podrobný přehled 12). Teprve s plně vyvinutou mentalizací je u člověka plně vyvinuto sebeuvědomování.
Poznámka
Výpočet dědivosti, neboli heritability, není jednoduchý.
Viz http://en.wikipedia.org/wiki/Heritability
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Sources
1. Addis, D.R., Wong, A.T., Schacter, D.I. Remembering the past and imagining the future: common and distinct neural substrates during event construction and elaboration. Neuropsychologia 2007, 45, p. 1363-1377.
2. Buckner, R., Andrew-Hanna, J., Schacter, D. The brain’s default network: anatomy, function and relevance to disease. Ann. NY Acad. Sci. 2008, 1124, p. 1-38.
3. Fair, D.A. Cohen, A.L., Dosenbach, N.U.C. et al. The maturing architecture of the brain´s default network. Proc. Natl. Acad. Sci. USA 2008, 105, p. 4028-4032.
4. Fox, M.D., Snyder, A.Z., Vincent, J.L. et al. The human brain is intrinsically organized into dynamic anticorrelated functional network. Proc. Natl. Acad. Sci. USA 2005, 102, p. 9673-9678.
5. Glahn, D.C., Winkler, A.M., Kochunov, P. et al. Genetic control over the resting brain. Proc. Natl. Acad. Sci. USA 2010, 107, p. 1223-1228.
6. Greene, J.D., Sommerville, R.B., Nystrom, L.E. et al. An fMRI investigation of emotional engagement in moral judgement. Science 2001, 293, p. 2105-2108.
7. Hahn, B., Ross, T.J., Stein, E.A. Cingulate activation increases dynamically with response speed under stimulus unpredictability. Cereb. Cortex 2007, 17, p. 1664-1671.
8. Chiao, J.Y., Harada, T. Komeda, H. et al. Neural basis of individualistic and collectivistic views of self. Hum. Brain. Mapp. 2009, 30, p. 2813-2820.
9. Chiao, J.Y., Harada, T., Komeda, H. et al. Dynamic cultural influences on neural representatiton of self. J. Cogn. Neurosci. 2009, 21, p. 1-11.
10. James, W. Principles of psychology. New York: Henry Holt and Co., 1890. Dostupné na: http://ia341341.us.archive.org/1/items/theprinciplesofp01jameuoft/theprinciplesofp01jameuoft.pdf
11. Johnson, M.K., Raye, C.I., Mitchell, K.J. et al. Dissociating medial frontal and posterior cingulate activity during self-reflection. SCAN 2006, 1, p. 56-64.
12. Koukolík, F. Před úsvitem, po ránu. Eseje o dětech rodičích. Praha: Karolinum, 2008, s. 177.
13. Koukolík, F. Funkční systémy lidského mozku. Praha: Galén, 2011 (v tisku).
14. Leary, M.R., Buttermore, N.R. The evolution of human Self: tracing the natural history of self-awareness. J. Theory. Soc. Behav. 2003, 33, p. 365-404.
15. Li C - S.R., Yan, P., Berquist, K.L. et al. Greater activation of the „default“ brain regions precede stop signal errors. NeuroImage 2007, 38, p. 640-648.
16. Mason, M.F., Bortin, M.I., Van Horn, J.D. et al. Wandering minds: the default network and stimulus-independent thought. Science 2007, 315, p. 393-395.
17. Otti, A., Guendel, H., Laer, L. et al. I know the pain you feel - how the human brain’s default mode predicts our resonance to another’s suffering. Neuroscience 2010, 169, p. 143-148.
18. Panksepp, J., Northhoff, G. The trans-species core SELF: the emergence of aktive cultural and neuro-ecological agents through self-related processing within subcortical-cortical midline networks. Conscious. Cogn. 2009, 18, p. 193-215.
19. Prior, H., Schwarz, A., Güntürkün, O. Mirror-induced behavior in the Magpie (Pica ): Evidence of self-recognition. PLoS. Biol. 2008 6(8), e202. doi:10.1371/journal. pbio.0060202.
20. Raichle, M.E., McLeod, M., Snyder, A. et al. A default state of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, p. 676-682.
21. Saxe, R., Kahnwisher, N. People thinking about thinking people: the role of temporo-parietal function in „theory of mind“. NeuroImage 2003, 19, p. 1835-1842.
22. Sheline, Y.I., Barch, D.M., Price, J.L. et al. The default mode network and self-referential processes in depression. Proc. Natl. Acad. Sci. USA 2009, 106, p. 1942-1947.
23. Supekar, K., Musen, M., Menon, V. Development of large-scale functional brain networks in children. PLoS. Biol. 2009, 7(7), e1000157. doi:10.1371/journal.pbio.1000157.
24. Supekar, K., Uddin, L.Q., Prater, K. et al. Development of functional and structural connectivity within the default mode network in young children. NeuroImage 2010, 52, p. 290-301.
25. Vanhaudenhyuse, A., Noirhomme, Q., Tshibanda, L.T-F. et al. Default network connectivity reflects the level of concisousness in non-communicative brain-damaged patients. Brain 2010, 133, p. 161-171.
26. Vincent, J.L., Patel, H., Fox, M.D. et al. Intrinsic functional architecture in the anesthetized monkey brain. Nature 2007, 447, p. 83-86.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2010 Issue 10
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
- Metamizole vs. Tramadol in Postoperative Analgesia
Most read in this issue
- Dušnost ve stáří podmíněná onemocněním plic
- Burn-out syndrom u lékařů
- Současný pohled na diagnostiku a léčbu karcinomu slinivky břišní
- Celoživotní vzdělávání všeobecných praktických lékařů v ČR a e-learning