
Evoluce a evoluční teorie pro lékaře (VII. Speciace neboli vznik nových druhů)
Evolution and evolutionary theory for physicians. VII. Speciation or rise of new biological species
Speciation is a term that denotes evolutionary processes by which new biological species arise. The concept of “species” is very broad and has a lot of meanings. Speciation is divided into allopatric (dichopatric and peripatric), parapatric and sympatric, as well as common hybrid speciation. The Wallace effect is a mechanism whereby natural selection strengthens reproduction isolation. Current research is focused on the genetic basis of speciation. Four genes have been described whose function forms the basis for hybrid sterility. The rate of speciation is discussed. The theory of punctuated equilibrium is at odds with the theory of phyletic gradualism.
Key words:
evolution, speciation, species concept, reproduction isolation.
Authors:
F. Koukolík
Authors‘ workplace:
Oddělení patologie a molekulární medicíny, Národní referenční laboratoř prionových chorob, Fakultní Thomayerova nemocnice s poliklinikou, Praha, Primář: MUDr. František Koukolík, DrSc.
Published in:
Prakt. Lék. 2010; 90(7): 391-395
Category:
Editorial
Overview
Speciace je soubor evolučních procesů, jimž vznikají nové biologické druhy. Pojem druhu je velmi široký a má mnoho významů. Rozlišuje se speciace alopatrická, (dichopatrická a peripatrická), parapatrická a sympatrická, častá je hybridní speciace. Wallaceův efekt je mechanismus, jímž přírodní výběr posiluje reprodukční izolaci. Současný výzkum se zaměřuje na genetické podklady speciace. Byly popsány geny, jejichž funkce je podkladem sterility hybridů. Rychlost speciace je předmětem diskuse. V rozporu s teorií fyletického gradualismu je teorie přerušované rovnováhy.
Klíčová slova:
evoluce, speciace, pojem druhu, reprodukční izolace.
Úvod
Počet žijících druhů života na Zemi se odhaduje v rozmezí 5–100 milionů, popsány jsou přibližně dva miliony (20). Jestliže jsou všechny současné podoby života potomkem posledního univerzálního společného předka, pak se objevuje otázka kdy, jak a proč se v čase a prostoru objevil tak velký a rozmanitý počet různých druhů života – jinak řečeno, co je podkladem biodiverzity.
Charakteristika druhu
Druh (species) je pojem, který má podle Mayra (13) tři základní významy:
Druh jako pojem:
typologický pojem druhu byl koncem 19. a v prvních desetiletích 20. století nahrazen biologickým pojmem druhu. Typologický pojem druhu neodpovídal druhům coby populacím a jejich evolučnímu potenciálu. Prubířským kamenem biologického pojetí druhu je zejména reprodukční kompatibilita – možnost mít plodné potomky (12, 13).
Druh jako taxon:
studium druhů v geografickém prostoru ukazuje, že jsou tvořeny početnými lokálními populacemi, které se vzájemně méně nebo více liší. Soubor populací distribuovaný v geografickém prostoru je druh jako taxon. Druh jako taxon je mnohorozměrný, druh jako pojem je bezrozměrný. Druhy jako taxony, které mají dobře definované součásti, poddruhy, se označují jako polytypické druhy.
Druh jako kategorie:
je příčka linnéovské hierarchie udělená taxonu považovanému za druh.
Vyčerpávající přehled současných definic pojmu species je v tab. 1, str. 395.
Speciace, aneb nové biologické druhy
Speciace
je pojem označující soubor evolučních procesů, jimiž vznikají nové biologické druhy (17). V průběhu těchto jevů je nutné odlišovat anagenezu od fylogeneze.
Anageneze, též „fyletická změna“
je pohyb „vzhůru“ na fylogenetické linii, postupná změna z původní do odvozené podoby života. Na konci anagenetického procesu se celá populace odlišuje od populace rodičovské, ta se považuje za vyhynulou.
Fylogeneze
označuje naproti tomu štěpení evolučních linií, vznik nových větví, neboli kladů fylogenetického stromu, který je spíše než jediným kmenem s mnoha větvemi vzájemně propojeným houštím (13, 14, 9,10).
Rozlišuje se několik typů speciace:
v průběhu alopatrické speciace
se populace štěpí na dvě izolované populace. Jestliže se po nějaké době takové populace opět setkají, jsou natolik odlišné, že nejsou s to vyměňovat geny – jsou tedy reprodukčně izolované. Existují dvě podoby alopatrické speciace: dichopatrická a peripatrická (13, (obr. 1).
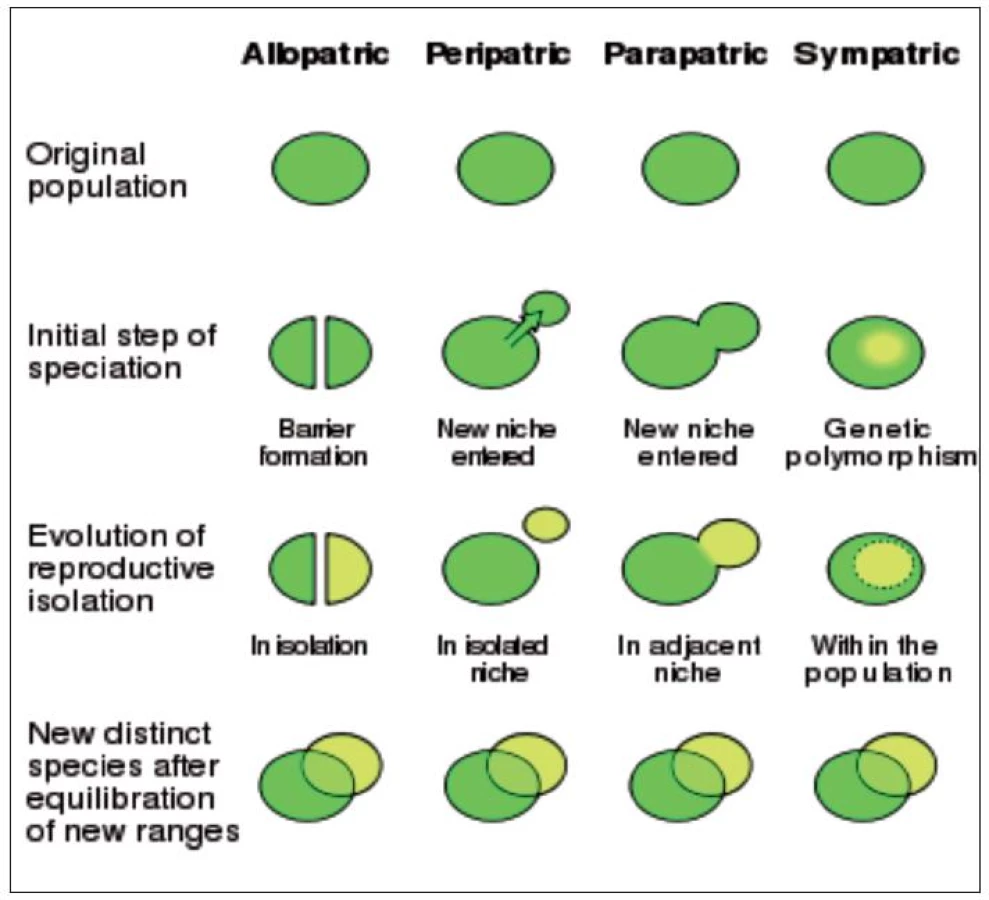
Při dichopatrické speciaci
je izolace důsledkem vzniku geografické bariéry, která populaci rozdělí. Příkladem je vznik Behringovy úžiny na konci pleistocénu, která vytvořila barieru mezi Sibiří a Aljaškou, což spustilo speciaci původně jednotných holoarktických druhů. Jiným příkladem je vznik nových druhů v ostrovech tropického dešťového pralesa, které se uchovaly v průběhu suchých období pleistocénu. Příkladem alopatrické speciace jsou některé ostrovní živočišné druhy.
Při peripatrické speciaci
se uplatňuje zakladatelská populace, která je za periferií populace rodičovské. Taková populace může stejně zaniknout, jako se může vyvinout do nového druhu. Zakladatelská populace je malá, proto geneticky ochuzená, může ji tvořit jen několik jedinců případně i jediná oplodněná samička. Genofond zakladatelské populace je proto podstatně chudší, než je genofond populace rodičovské. Zakladatelská populace je vystavena vyššímu selekčnímu tlaku v novém prostředí. Příkladem peripatrické speciace je moskyt Culex pipiens, který osídlil londýnské metro v průběhu 19. století. Od populace žijící na povrchu se liší geneticky i chováním (2).
Při parapatrické speciaci,
to je další podoba speciace, vstupuje část populace do nové niky, v ní se vyvine reprodukční izolace, zóny obývané oběma populacemi jsou však částečně společné. Příkladem jsou racci Larus tvořící druhový prstenec kolem severního zemského pólu.
Sympatrická speciace
je označení vzniku dvou nebo většího počtu populací obývajících společné území a přitom pocházejících z jedné populace rodičovské. Existence sympatrické speciace je předmětem diskuse. Jako příklad sympatické speciace se uvádí Rhagoletis pomonella (apple maggot fly, hawthorn fly, vrtule jabloňová) jejíž různé populace se živí různými plody. V severní Americe se objevila v průběhu 19. století populace, která se začala živit jablky, tedy druhem, jenž byl dopraven ze zámoří, nikoli plody hlohu, jímž se rodičovská populace Rhagoletis (převážně) živila do té doby. Od původní populace se Rhagoletis pomonella liší geneticky i chováním, hybridizace s rodičovskou populací je nízká. Rhagoletis pomonella se někdy uvádí jako příklad rychlé evoluce „před očima“, podobně jako speciace koljušek, cichlid nebo proslulých Darwinových pěnkav na Galapágách.
Diskutovanou vzácností je speciace podmíněná vzdáleností, neboli cirkulární překrývání. Dlouhý řetěz populací vyústí v překrytí východiska a konce. Prokazuje se u racků Larus argentatus a mloka Ensantina (13).
Wallaceův efekt
je mechanismus, jímž přírodní výběr posiluje reprodukční izolaci (17). Může se objevit ve chvíli, kdy se dvě rozdělené populace téhož druhu po nějaké době setkají. Jestliže byla jejich reprodukční izolace úplná, jsou z nich dva odlišné druhy. Jestliže byla neúplná, může jejich páření vést ke vzniku hybridů, kteří mohou být jak plodní, tak neplodní.
Jestli jsou jejich hybridy neplodní, nebo je jejich biologická zdatnost (fitness) nižší než u jejich předků, další reprodukční izolace nenásleduje. Příkladem jsou hybridi koní a oslů, muly a mezci.
Jestliže jsou hybridi biologicky zdatnější než jejich rodičovské populace, smísí se zpětně s populací toho druhu s nímž jsou v kontaktu.
Wallaceův efekt v literatuře označovaný jako reinforcement (v tomto případě doslovně zesílení reprodukční izolace) se považuje za podklad jak parapatrické, tak sympatrické speciace. Bez Wallaceova efektu by se oblast kontaktu mezi odlišnými podobami téhož druhu, této oblasti se říká hybridní zóna nebo hybridní pás, na hranici mezi odlišnými druhy neproměnila.
Velmi zajímavý je v této souvislosti objev několika procent genů jaderné DNA neandrtálců v genomu současné populace Homo sapiens, zatímco mitochondriální geny neandrtálců – ty se dědí jen po matkách – součástí genomu současného člověka nejsou Neandrtálci se totiž po oddělení od linie směřující od společného předka k Homo sapiens znovu s Homo sapiens setkali – oddělení přitom trvalo několik set tisíc let (1, 5).
Izolační mechanismy
Izolační mechanismy jsou biologické vlastnosti jednotlivých organismů blokující křížení populací různých druhů tam, kde obývají společné území.
Existují prezygotickémechanismy izolace blokující křížení druhů: možní partneři křížení se vůbec nesetkají, protože je izoluje roční doba nebo habitat, dále odlišnosti chování, čemuž se říká etologická izolace, případně jsou izolováni mechanicky: při kopulaci nedojde k přenosu spermatu.
Mezi postzygotickémechanismy náleží
- a) inkompatibilita gamet – spermie neoplodní vajíčko,
- b) zygotická mortalita při níž oplodněné vajíčko zanikne,
- c) vznikne hybrid s omezenou životností,
- d) vznikne hybrid, který životný je, ale je částečně nebo úplně sterilní, případně plodí defektní potomstvo.
Příklady speciace
Tradičně uváděným příkladem přirozené speciace je tříostná koljuška (Gasterosteus aculeatus, (obr. 2). Odhaduje se, že soudobá druhová rozmanitost tříostných koljušek vyžadovala od konce poslední ledové doby asi 10 000 generací a projevuje se odlišnostmi v zabarvení, uspořádání ploutví i kostry včetně čelistí.

Hendry et al. (6) na příkladu speciace koljušek dokládá jak je speciace složitý proces. Speciaci považuje za kontinuum, v němž je možné rozlišovat několik stavů:
- trvalé variace v populacích schopných vzájemného míšení, bez genetického nebo behaviorálního omezení – všichni jedinci jsou potenciálními partnery,
- částečně diskontinuální variace s menšími reprodukčními izolacemi,
- silně diskontinuální variace s výraznou, nicméně reverzibilní reprodukční izolací,
- kompletní a ireverzibilní reprodukční izolace.
Výzkum koljušek ukazuje jak proměny uvnitř jednotlivých stavů, tak přechody mezi stavy. Většina populací je ve stavu 1, nicméně i v těchto populacích je možné doložit selekci a výběrové páření. Přechod do stavu 2 bývá spjat s geografickou izolací doprovázenou parapatrií (například rozdílem života v jezerech a v proudící vodě) nebo alopatrií (rozdílem života v různých jezerech). Přechod do stadia 3 se objevuje, jakmile se dostanou parapatrické nebo alopatrické populace, které se vyvíjely díky silnému divergentnímu výběru, do druhotného kontaktu, objevuje se však i v parapatrických a alopatrických populacích. Stav 4 je charakterizován změnou chromozomálního uspořádání a genetickou inkompatibilitou nezávislou na prostředí – příkladem jsou reprodukčně zcela nebo téměř zcela izolované koljušky žijící v Japonském moři a koljušky žijící v Tichém oceánu. V průběhu speciace se tedy uplatňuje řada mechanismů:
- selekce,
- adaptace,
- mutace, i
- geografické vlivy.
Vztah genů a ekologických vlivů při speciaci koljušek, původně mořských ryb, které se opakovaně adaptovaly na sladkovodní prostředí, popisují Schluter a Conte (19). Mají za to, že se ekologicky podmíněná speciace odehrávala opakovaně a paralelně. Selekce podmíněná sladkovodním prostředím působila na variace uchovávané v mořských populacích „exportem“ alel adaptovaných na sladkou vodu z míst, které byly s mořskými populacemi v kontaktu.
Jiným oblíbeným modelem dramaticky rychlé speciace a adaptivní radiace jsou africké cichlidy (vrubozubcovití). Tato čeleď ryb obnáší více než 1 000 velikostí, tvarem, barvou i chováním nesmírně rozmanitých (a krásných) druhů v tropických a subtropických oblastech tří světadílů. V jezerech východní Afriky byly objeveny stovky endemických druhů, které se velmi rychle vyvíjely díky přírodnímu i pohlavnímu výběru (obr. 3) (18)

Hybridizace
Hybridní speciace
je označení teorie, podle níž má zásadní význam při vzniku nového druhu hybridizace. Je častější, než se dosud mělo za to, a to jak u rostlin, tak u živočichů (11). Označení se týká hybridních druhů, které zdvojily počet svých chromozomů, tomu se říká allopolyploidie. Odvozený druh původně vlastní celý genom každého ze svých rodičů, vklad každého rodiče je 50%, což nevylučuje následnou rekombinaci a genovou konverzi, takže u starších polyploidních druhů může být vklad rodičů nerovný.
Mallett (11) v přehledném článku uvádí, že s jiným druhem hybridizuje přibližně 10 % živočichů a 25 % rostlin, přičemž je hybridizace vysoká u rychle se šířících druhů. Příkladem je 75% hybridizace britských kachen (Anatideae).
Vztah speciace a přírodního výběru
Na vztah speciace a přírodního výběru uvádí Via (21) dva odlišné pohledy. Prvnímu, klasickému, říká „pohled dalekohledem“. Druhý, současný, pojmenovala „pohled lupou“.
Představitelé „pohledu dalekohledem“ jsou zejména Ernst Mayr a Theodosius Dobzhansky. Mayr v r. 1942 vytvořil biologický pojem druhu (tab. 1), těžištěm speciace je podle tohoto autora reprodukční izolace. Mayr zdůrazňoval, že se reprodukční izolace může uplatnit jen tam, kde geografická izolace zamezí toku genů, výsledkem je alopatrická speciace. Dobzhansky se koncem třicátých let minulého století, podobně jako H. J. Mueller na začátku let čtyřicátých, soustředil zejména na mechanismy postzygotické izolace.

Batesonův-Dobhzhanského-Muellerův model (23) vysvětloval, jak se může vyvinout sterilita hybridů, aniž by selekce zastavila intermediární články procesu. V současnosti byl tento model vzat v potaz (7). „Pohled dalekohledem“ je retrospektivní analýza, odtud pojmenování, prováděná na úrovni druhů.
Model pojmenovaný „pohled lupou“ je analýza ekologických a genetických příčin reprodukční izolace na úrovni populací. Tento pohled předpokládá, že se při divergentní selekci sympatrických populací stávají genomy nově vznikajících druhů dočasnou genovou mozaikou. Ekologicky významné genové oblasti v takové mozaice odolávají výměně i při zachovaném genovém toku.
Genetické podklady speciace
Postzygotická isolace je
- extrinsická, a
- instrinsická (23).
Extrinsická izolace
je podmíněna destruktivními důsledky interakce hybridu s prostředím: hybridi jsou geneticky vybaveni pro život v nikách rodičovských prostředí, které neodpovídají jejich nice.
Instrinsická postzygotická izolace
je způsobena vývojovými defekty hybridů, jejichž výsledkem je neživotnost nebo částečná či úplná sterilita. Uvádí se, že exstrinsická izolace je častější a významnější než intrinsická zejména v raných fázích druhové divergence. Podklady intrinsické postzygotické isolace jsou:
- úroveň ploidie,
- přeskupování chromozomů doprovázené novým uspořádáním (rearrangement),
- genová inkompatibilita, a
- interakce mezi jaderným genomem a endosymbiontem, což se oprávněně považuje za zvláštní případ předchozího mechanismu (8). Za nejvýznamnější mechanismus se považuje genová inkompatibilita.
U živočichů byly dosud popsány čtyři geny, které jsou podkladem sterility hybridů a to:
- Odysseus,
- JYalfa,
- Overdrive u octomilek, a
- Prdm9 (Meisetz) u myší.
Myší protein Prdm9 se vyvíjel s vysokou rychlostí v doméně, která se váže na DNA a obsahuje „zinc fingers“ (doslovně zinkové prsty), což jsou malé proteinové motivy užívající jeden nebo víc atomů zinku ke stabilizaci prostorového uspořádání. Lze je nejlépe chápat jako interaktivní moduly vážící molekuly DNA, RNA, proteiny. Odlišnost jediného z nich rozlišuje ty alely genu Prdm9, které jsou příčinou sterility hybridů, od alel, které příčinou sterility nejsou.
Oliver et al. (15) ukázali, jak pozitivní selekce rychle změnila počet a sekvence Prdm9 „zinkových prstů“ ve 13 genomech hlodavců. Typ pozitivní selekce Prdm9 „zinkových prstů“ dokládá, jak působila rychlá evoluce na rozhraní mezi Prdm9 proteinem a DNA sekvencemi, na které se váže. Podobný stav byl doložen u dalších taxonů rozlišných živočichů včetně primátů. Variabilita alel lidských Prdm9 „zinkových prstů“ je v asociaci se sníženým rizikem neplodnosti. Prdm9 tedy ovlivňuje mezidruhovou sterilitu u myší a vnitrodruhovou sterilitu u lidí. Opakované epizody positivní selekce Prdm9 ukazují, že se spolu s ním rychle vyvíjely sekvence DNA, na které se váže. Podle Olivera et al. (15) je molekulárním podkladem sterility zprostředkovávané genem Prdm9 neslučitelnost mezi vazebnou specificitou proteinu Prdm9 a satelitní DNA.
Qvarnström a Bailey (16) uvádějí v přehledném článku o speciaci podmíněné evolucí genů vázaných na pohlavní chromozomy čtyři rozdíly mezi pohlavními chromozomy a autozomy:
- relativní rychlost evoluce,
- nenáhodnou akumulaci genů,
- výskyt inkompatibilních recesivních genů u hybridů, a
- podílem rekombinací.
V raných stádiích rozdělování populace jsou rozdíly dány autozomálními geny. Rychle se vyvíjející geny vázané na pohlavní chromozomy mají zřejmě klíčový význam pro vývoj sexuální izolace tím, že kódují ovlivňující biologickou zdatnost (fitness) vázanou na pohlaví, například primární a sekundární pohlaví znaky. Geny kódující znaky, které jsou podkladem sexuální izolace, jsou s vyšší pravděpodobností příčinou sterility hybridů, jsou-li vázány na pohlavní chromozomy.
Rychlost speciace
Rychlost speciace je předmětem diskuse. Někteří evolucionisté mají za to, že speciace běží konstantně, pomalu, v průběhu dlouhých geologických obdobích, v tomto případě se mluví o fyletickém gradualismu (12).
Naproti tomu Niles Eldredge a Stephen J. Gould (3, 4), představitelé teorie přerušované rovnováhy (punctuated equilibrium), dokazují, že jsou druhy dlouhou dobu stálé a speciace proběhne v poměrně krátkém období – jejich teorie je tedy s fyletickým gradualismem v rozporu.
MUDr. František Koukolík, DrSc.
Oddělení
patologie a molekulární medicíny
Národní referenční
laboratoř prionových chorob
Fakultní Thomayerova
nemocnice s poliklinikou
Vídeňská 800
140 59 Praha 4 Krč
E-mail:
frantisek.koukolik@ftn.cz
Sources
1. Burbano, H.A., Hodges, E., Green R.E. et al. Targeted investigation of the neandertal genome by array-based sequence capture. Science 2010, 328, p. 723-725.
2. Byrne, B., Nichols, R.A. Culex pipiens in London underground tunnels: differentiation between surface and subterranean populations. Heredity 1999, 82, p. 7-15.
3. Eldredge, N., Gould, S.J. Punctuated equilibria: an alternative to phyletic gradualism. In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper 1973. p. 82-115. Reprinted in N. Eldredge Time frames. Princeton: Princeton Univ. Press, 1985.
4. Gould, S.J., Eldredge, N. Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology 1977, 3, p. 115-151.
5. Green, R.E., Krause, J., Briggs, A.W. et al. A draft sequence of the neandertal genome. Science 2010, 328, p. 710-722.
6. Hendry, A.P., Bolnick, D.I., Berner, D. et al. Along the speciation continuum in sticklebacks. J. Fish. Biol. 2009, 75, p. 2000-2036.
7. Gourbiére, S., Mallet, J. Are species real? The shape of the species boundary with exponential failure, reinforcement, and the ‘missing snowball’. Evolution 2010, 64, p. 1–24.
8. Hoffmann, A.A., Rieseberg, L.H. Revisiting the impact od inversion in evolution: from population genetic markers to drivers of adaptive shift and speciation? Annu. Rev. Ecol. Evolv. Syst. 2008, 39, p. 21-42.
9. Koonin, E.V., Wolf, Y.I. Genomics of bacteria and archaea: the emerging generalizations after 13 years. Nucleic. Acid. Res. 2008, 36, p. 6688-6719.
10. Koonin, E.V. Darwinian evolution in the light of genomics. Nucleic. Acid. Res. 2009, 37, p. 1011-1034.
11. Mallett, J. Hybrid speciation. Nature 2007, 446, p. 279-283.
12. Mayr, E. Change of genetic environment and evolution . In J. Huxley, A. C. Hardy and E. B. Ford. Evolution as a Process. London: Allen and Unwin, 1954, p. 157-180.
13. Mayr, E. What evolution is. New York: Basic Books, 2001.
14. Mueller, G.B. Evo-devo as a discipline. In: Evolving pathways: key themes in evolutionary developmental biology. Minelli A, Fusco G. (eds.) Cambridge: Cambridge University Press 2008, p. 3-29.
15. Oliver, P.L., Goodstadt, L., Bayes, J.J. et al. Accelerated evolution of the Prdm9 speciation gene across diverse metazoan taxa. PLoS. Genet, 5(12), e1000753. doi:10.1371/journal.pgen.1000753
16. Qvarnström, A., Bailey, R.I. Speciation through evolution of sex-linked genes. Heredity 2009, 102, p. 4-15.
17. Ridley, M. Speciation - What are the major theories of speciation? [on-line]. Dostupné na http://www.blackwellpublishing.com/ridley/tutorials/Speciation6.asp.
18. Salzburger, W. The interaction of sexually and naturally selected traits in the adaptive radiation of cichlid fishes. Mol. Ecol. 2008, 18, p. 169-185.
19. Schluter, D., Conte, G.L. Genetics and ecological speciation. Proc. Natl. Acad. Sci. USA 2009, 106, suppl 1, p. 9955-9962.
20. The Tree of Life Web Project (ToL) [on-line]. Dostupný na http://tolweb.org/tree/.
21. Via, S. Natural selection in action during speciation. Proc. Natl. Acad. Sci. 2009, 106, Suppl 1, p. 9939-9946.
22. Wilkins, J.S. (2006) A list of 26 species concepts [on-line]. Dostupné na http:// scienceblogs.com/evolvingthoughts/2006/10/a_list_of_26_species_concepts.php.
23. Wolf, J.B.W., Lindell, J., Backström, N. Speciation genetics: current status and evolving approaches. Phil. Trans. R. Soc. B 2010, 365, p. 1717-1733.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2010 Issue 7
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
Most read in this issue
- Operace šilhání u dospělých – ano, nebo ne?
- Transplantace jater u dětí
- Fibrilace síní a elektrická kardioverze v kardiologické ambulanci
- Asplenický pacient: odpovědnost nás všech