
Základy kognitivní, afektivní a sociální neurovědy XXV. O štěstí – hédonický mozek
Basics of social cognitive and affective neuroscience XXV. On happiness – the hedonic brain
Ever since Aristotle, happiness has been thought of as pleasure (hedonia) and a life well-lived (eudamonia). Substantial progress in understanding the neurobiology of sensory pleasure has been made. Interspecies pleasure research says that pleasure manifests in consciousness (subjective liking) and in brain and behavioural reactions (objective liking). Beyond the feeling of liking there are components of reward that contain wanting, motivation for reward and learning, associations, representations and predictions about future rewards. The “hedonic brain” is a description of the neuroanatomy of pleasure, a large scale neuronal network consisting of subcortical nuclei, eg. periaquaductal gray, ventral tegmental area, ventral palidum and nc. accumbens (ventral striatum) and of cortical parts, orbitofrontal, cingulate, medial frontal and insular cortices. Pleasure, eudamonic happiness, may arise due to the function of brain’s default system.
Keywords:
reward – pleasure – hedonic brain
Authors:
F. Koukolík
Authors‘ workplace:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Thomayerova nemocnice, Praha
; Oddělení patologie a molekulární medicíny
Published in:
Prakt. Lék. 2013; 93(1): 10-15
Category:
Of different specialties
Overview
Od Aristotelových dob se uvažuje o slastné, neboli hédonické a eudaimonické stránce štěstí, tou je míněn „správně žitý život.“ Pochopení neurobiologie senzorické slasti učinilo podstatný krok dopředu. Mezidruhový výzkum zjistil, že se slast manifestuje ve vědomí (subjektivním pocitem „mám to rád“), dále objektivně v mozku a v chování („mám to rád“). Kromě pocitu „mám to rád“ jsou složkami odměny chtění, které je motivací pro zisk odměny, a učení, které je podkladem asociací, reprezentací a předpovědí týkajících se zisku příští odměny. „Hédonický mozek“ je popisem neuroanatomie slasti. Tvoří ho oboustranná neuronální síť velkého rozsahu s podkorovými oblastmi, např. šedí kolem mokovodu, area tegmentalis ventralis, ventrálním palidem a ventrálním striatem (nc. accumbens), jakož i části korové, orbitofrontální, cingulární, mediální prefrontální a insulární kůra. Smysl slasti, eudaimonický aspekt štěstí, může být výsledkem činnosti implicitního systému mozku.
Klíčová slova:
odměna – slast – hédonický mozek
Úvod
Od Aristotelových dob (384–322 př.n.l) se mluví o dvou aspektech štěstí, a to hédonickém a eudaimonickém. O pojmu hédonia říkají slovníková vymezení, že vystihuje vědomý pocit něčeho příjemného a je afektivní složkou štěstí. Eudamónia je pojem, který lze chápat jako rozumovou stránku štěstí, obnáší správně a vyrovnaně žitý život. Třetím, modernějším aspektem štěstí je pocit podílu na životě, pocit „jsem součástí života“ (20). Hédonický a eudaimonický aspekt štěstí jdou obvykle ruku v ruce. V průzkumech, které se táží se na pocit štěstí, odpovídá kolem 80 % tázaných, že jejich eudamonická satisfakce je nadprůměrná. Stejný podíl tázaných říká, že je podobně nadprůměrné i jejich hédonické cítění (6–7 bodů z desetičlenné stupnice, v níž 5 bodů označuje hédonickou neutralitu (17). Je ovšem nutné uvážit možnost sebestylizace tázaných lidí.
Současné metody popisují strukturu a funkci neuronálních sítí velkého rozsahu, jejichž činnost je podkladem hédonické zkušenosti, tedy „hédonický mozek“ (21), o něco lépe než těch sítí, jejichž činnost je podkladem zkušenosti eudaimonické.
Vývoj
U myší, krys, opic, šimpanzů, orangutanů i novorozených dětí je možné vložením něčeho sladkého do úst vyvolat obličejovou reakci „mám to rád“ (liking). Projeví se vypláznutím jazyka, případně olizováním rtů. Opačnou, tedy averzivní reakci lze vyvolat vložením něčeho intenzivně hořkého. Reakci „mám to rád“ lze považovat za slastnou složku fundamentální odměny, jejímž příkladem je slastný pocit v průběhu přijímání potravy nebo v průběhu sexuálního chování. Tato složka může být sdílena se slastmi „vyššího řádu“, například v průběhu uměleckého prožitku hudebního, výtvarného nebo dramatického, stejně jako v průběhu altruistické nebo transcendentní zkušenosti. Kromě složky „mám to rád“ má fundamentální odměna složky další, které hédonické nejsou, a to složku „chci to“ (wanting), která je chápána jako motivace k získání odměny, a složku učení (learning) danou asociacemi, reprezentacemi a předpovědí budoucích odměn založenými na minulé zkušenosti (20, 21, 26–28). Jak pozitivní, tak negativní afekt lze chápat jako adaptivní reakce, což si uvědomil už Charles Darwin – proto jejich evoluční konzervace.
Nutné je odlišovat afektivní stavy, které lze objektivně měřit na základě fyziologických změn, dále změn chování i proměn činnosti mozku od vědomého afektivního pocitu, jenž je subjektivní stránkou emocí. Damasio (9, 10) odlišuje v tomto smyslu emoce, soubory nevědomých procesů, od pocitů, které považuje za vědomou emoční zkušenost.
Neuronální systémy
Slast lze tedy rozložit do tří složek: motivace („chci to“ – wanting), obliby („mám to rád“ – liking) a učení (learning). Každá z těchto složek má vědomou, neboli explicitní složku, a složku nevědomou, neboli implicitní. Explicitní složky prožíváme vědomě, například si uvědomujeme svou slast, touhu, očekávání nebo štěstí. Nevědomé, implicitní složky zpracovávají informace mimo dosah vědomé pozornosti. Mohou dodávat váhu, resp. význam podnětu, po kterém toužíme.
Uzly neuronální sítě velkého rozsahu, jejíž stavba a funkce je podkladem tvorby a prožívání slasti, neboli hédonických stavů, je na obrázku 1. Objem spouštěčů (hotspots) hédonické reakce v mozku hlodavců obnáší několik kubických milimetrů, v lidském mozku nanejvýš několik kubických centimetrů; jsou co do počtu poměrně vzácné. Spouštěče tvoří integrovanou síť (26) a byly zjištěny v plášti nc. accumbens, ventrálním palidu, pravděpodobně existují i v dalších oblastech mozku, například v parabrachiálním jádře mozkového kmene a dalších limbických a telencefalických oblastech.

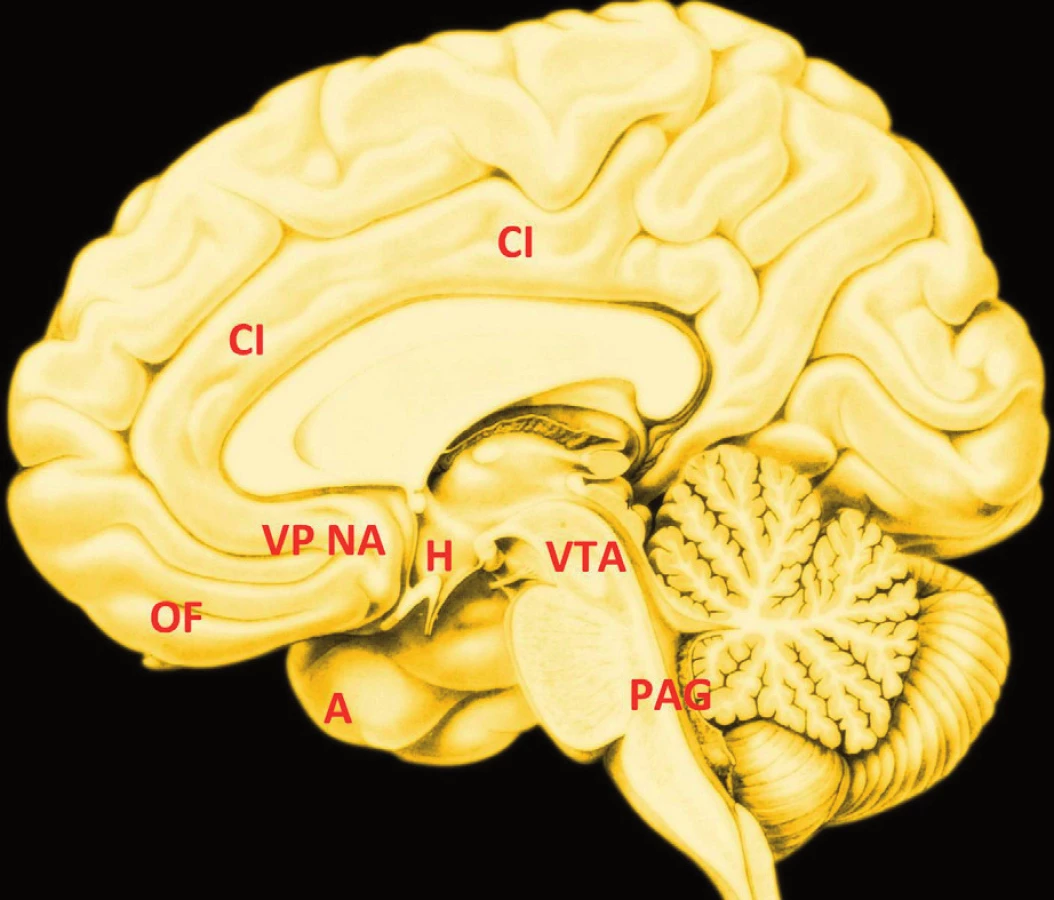
Jestliže se spouštěče ovlivní opioidními, endokanabiodními nebo dalšími chemickými modulátory (to lze docílit v experimentu mikroinjekcí), zvýší reakci „mám to rád“. Slast vyvolaná spouštěči se stává podkladem motivace tím, že spustí systém „chci to“ (wanting). Tento systém přikládá podnětům přitažlivost, jakmile je jim přiřazen mezolimbickým systémem (3). Oba systémy mohou být činné nezávisle, takže se stává, že člověk něco (nutkavě) chce, aniž by to měl rád. Předpokládá se, že právě tato skutečnost může být podkladem chemické závislosti v rozvinuté fázi: Závislí lidé nutkavě chtějí alkohol, případně jiné látky, aniž by jim tyto látky poskytovaly nějakou slast. Mechanismus „chci to“ je převážně podkorový, neovlivnitelný vědomou pozorností a není závislý na vědomé paměti a vědomém plánování. Chemicky závislí lidé proto dobře vědí, že jim látka, kterou zneužívají, neprospívá, přesto ji nutkavě vyžadují.
Jestliže je slast „nátěrem“ jakéhokoliv signálu, který může být příjemný (12), přičemž různým lidem a zvířatům mohou být příjemné i protikladně rozličné signály, pak platí otázka, jakým způsobem je tento „nátěr“ kódován do řeči neuronálních systémů mozku.
Ventrální palidum a nucleus accumbens
Ventrální palidum je ventrální částí globus palidus, součásti bazálních ganglií, systému, jehož dalšími částmi jsou nc. caudatus, putamen (společně zvané neostriatum), entopedunkulární jádro, nc. subthalamicus a s. nigra. Tradiční představu bazálních ganglií coby systému sloužícího motorice rozšířilo současné poznání o podíl na kognitivních funkcích, emotivitě, motivaci a odměně (obr. 2) (16). Experimenty na hlodavcích doložily existenci hédonického spouštěče v zadní polovině ventrálního palida, podobně jako ho doložily v mediální části obalu nc. accumbens (někdy zvaného ventrální striatum), který s ventrálním palidem topograficky sousedí.
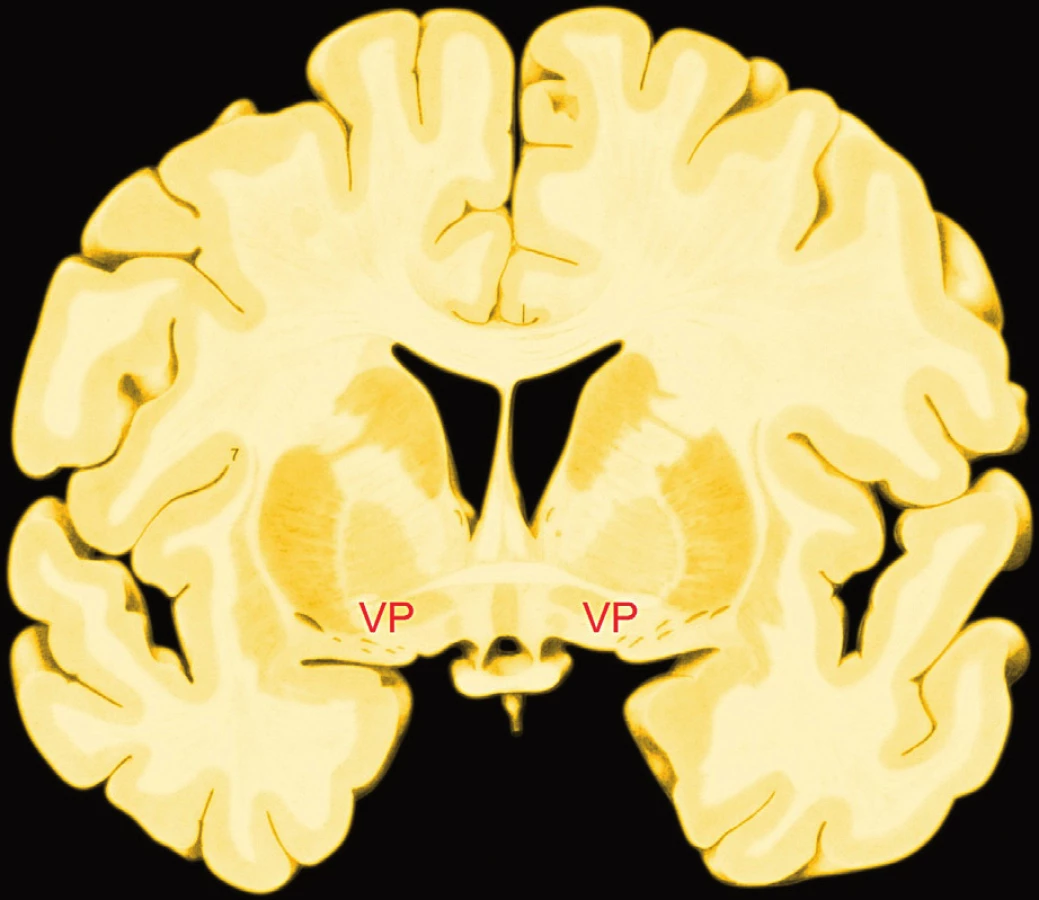
Opioidní stimulace spouštěče ve ventrálním palidu zdvojuje míru odpovědi „mám to rád“. Neurony této oblasti bouřlivě zvyšují aktivitu, jakmile pokusné zvíře ochutná cukr. Úměrně učení se následně činnost těchto neuronů stane odpovědí na podněty, které předpovídají sladkou odměnu. Přetrvávající hédonickou odpověď však kódují stále. Destrukce této oblasti nahradí odpověď „mám to rád“ odpovědí „mám to nerad“. Ventrální palidum u lidí odpovídá zvýšením aktivity na nejrůznější podněty chápané jako odměna, od potravy k penězům, a to i tehdy, jsou-li tyto podněty senzoricky podprahové, takže si je lidé neuvědomují, jak prokázal kromě jiných experiment s podprahovou stimulací peněžním podnětem (24) (tab. 1).

Ventrální palidum je cílem projekce z ventrálního striata (nc. accumbens), amygdaly, orbitofrontální a inzulární kůry, ventrálního tegmenta a nc. parabrachialis, což jsou jak podkorové, tak korové uzly hédonického mozku. Ventrální palidum projikuje cestou dorzomediálního thalamu do limbických částí prefrontální kůry – kůry orbitofrontální, cingulární a inzulární.
Miller et al. (23) popsal anhedonii, která byla důsledkem selektivního oboustranného poškození globus palidus u 34letého muže, v jehož anamnéze bylo chronické zneužívání alkoholu, LSD a dalších halucinogenů, závislost na marihuaně, kokainu, opiátech a extázi. Pít alkohol začal tento pacient ve svých 9 letech, s marihuanou začal ve 12, kokainem ve 13, s opiáty ve 20, s extází v 21 letech. Po předávkování metadonem proběhla epizoda hypotenze a hypoxémie, byl resuscitován. Kromě dalších neuropsychiatrických příznaků následovala deprese, anhedonie, potíže se soustředěním. Touha po drogách vymizela, začal abstinovat, pil jen sklenku vína příležitostně s večeří, víno mu nedělalo žádnou slast. Vyšetření magnetickou rezonancí našlo malou selektivní destrukci pars interna g. palidus vpravo a obou součástí g. palidus vlevo, aniž by byly poškozeny jiné části mozku. Kognitivní a řídící funkce zůstaly nedotčené.
Senzorickou odpověď (např. sladký vs. slaný) lze experimentálně oddělit od odpovědi hédonické („mám to rád“ vs. „mám to nerad“) i od mechanismu učení (27). Je pravděpodobné, že ventrální palidum kóduje i další dvě součásti slasti: „chci to“ a učení (1).
Korové oblasti
Korovými oblastmi hédonického mozku jsou kůra orbitofrontální, inzulární, mediální prefrontální a cingulární (obr. 1). Je nutné rozlišovat kódování a kauzalitu: Na kódování slasti lze soudit z korelace aktivity mozkové oblasti ve vztahu k příjemnému podnětu. Na kauzalitu, tedy možnost, že je činnost dané oblasti „příčinou“ pocitu slasti lze soudit z důsledků manipulace oblastí, například z důsledků stimulace či poškození.
Kódování slasti je dáno systémem, jehož vrcholem je střední a přední podoblast orbitofrontální kůry. Tato oblast mohutně odpovídá na slast z příjmu potravy (18), orgasmu (13), metamfetaminu (29) a poslechu hudby (5).
Kringelbach et al. (18) doložili významnou korelaci mezi aktivací zmíněného segmentu orbitofrontální kůry a příjmem příjemné tekuté potravy. Jakmile byl dosažen pocit sytosti, aktivace v obraze funkční magnetické resonance (fMR) klesla. Pokles aktivity byl ve vztahu k druhu přijaté potravy specifický.
Small et al. (25) testovali pozitronovou emisní tomografií (PET) aktivitu mozku dobrovolníků, kteří jedli čokoládu až do přesycení. Po každém kousku čokolády pokusné osoby oznámily, v jaké míře jim byla čokoláda příjemná nebo nepříjemná a zda chtějí další kousek. V době, kdy byla čokoláda příjemná, byla vysoce aktivní subkalosální ventromediální prefrontální kůra, zadní a vnitřní část orbitofrontální kůry, inzulární a přilehlá operkulární kůra, striatum a střední mozek. Jestliže dobrovolníci jedli čokoládu dál i v době, kdy již byli sytí, aktivovaly se parahipokampální závit, zevní a zadní očnicová kůra a prefrontální korové oblasti. Laterální a mediální očnicová kůra fungují zřejmě jako misky vah: Aktivace vnitřní části odpovídá příjemnému pocitu, aktivace zevní části odpovídá pocitu přesycení. Jak v průběhu kladného, tak v průběh záporného pocitu se aktivovala zadní cingulární kůra.
Georgiadis et al. (13) užili PET ke sledování aktivity mozku 12 zdravých žen v průběhu sexuální stimulace. Orgasmus koreloval kromě jiného s poklesem perfuze zvláště v levé zevní orbitofrontální kůře, dolním spánkovém závitu a v pólu spánkového laloku.
Völlmová et al. (29) vystavila zdravé, chemicky nezávislé dobrovolníky účinku metamfetaminu, vysoce adiktivní látky, která je agonistou dopaminergního systému. Činnost jejich mozku sledovala fMR. Amfetamin aktivoval mediální orbitofrontální kůru, rostrální část přední cingulární kůry a ventrální striatum (nc. accumbens) (obr. 1). Aktivace mediální orbitofrontální kůry byla nezávislá na motorické odpovědi. Již první podání amfetaminu tedy uvede do vysoké aktivity systém odměny. Aktivita orbitofrontální kůry se tedy podílí na posilování bludného kruhu závislosti na amfetaminu.
Bloodová a Zattore (5) ke studiu slasti plynoucí z poslechu hudby užili PET. Pokusné osoby si samy volily hudbu, o níž říkaly, že jim při jejím poslechu „běhá mráz po zádech“. U různých to byla různá hudba. Perfuze v mozku se v průběhu pocitu mrazení měnila ve ventrálním striatu, středním mozku, amygdale, orbitofrontální kůře a ventrální mediální prefrontální kůře. Slast plynoucí z poslechu hudby tedy aktivuje stejnou neuronální síť, jakou aktivují i ostatní slastné podněty.
Kromě ventrálního palida a ventrálního striata je tedy dalším klíčovým uzlem hédonického mozku orbitofrontální kůra (přehled 19). Je na ventrální ploše čelního laloku. Je definována projekcí velkobuněčné části mediálního jádra dorzomediálního thalamu, což ji odlišuje od jiných prefrontálních korových oblastí. Původní Brodmanova cytoarchitektonická mapa v této kůře rozlišovala tři oblasti: 10, 11 a 47. Současné členění je podstatně podrobnější (7). Na homologii orbitofrontální kůry lidí a ostatních primátů lze soudit ze zapojení a cytoarchitektury. Orbitofrontální kůra se od šestivrstvé dorzolaterální prefrontální kůry odlišuje jen pětivrstvou agranulární kůrou (ta je v jejích zadních a středních částech) a dysgranulární kůrou v části střední. Pouze v přední části je orbitofrontální kůra rovněž šestivrstevná. U lidí je orbitofrontální kůra variabilní, lze rozlišit tři typy. Kromě toho prodělává mezi 5. až 20. rokem věku intenzivní vývoj.
Orbitofrontální kůra je součástí funkčního systému OMPFC, tvořeného orbitofrontální kůrou, mediální prefrontální kůrou a přední cingulární kůrou. Orbitální část, která dostává vstupy ze všech smyslových modalit včetně viscerálních aferent, odpovídá Brodmanovým areám 11, 13 47/12 (obr. 3). Předpokládá se, že reguluje příjem jídla. Mediální části odpovídající area 11, 13, 14, laterální části area 47/12 a area 25, 32 a 10 na vnitřní ploše hemisféry jsou zdrojem mohutných visceromotorických výstupů.
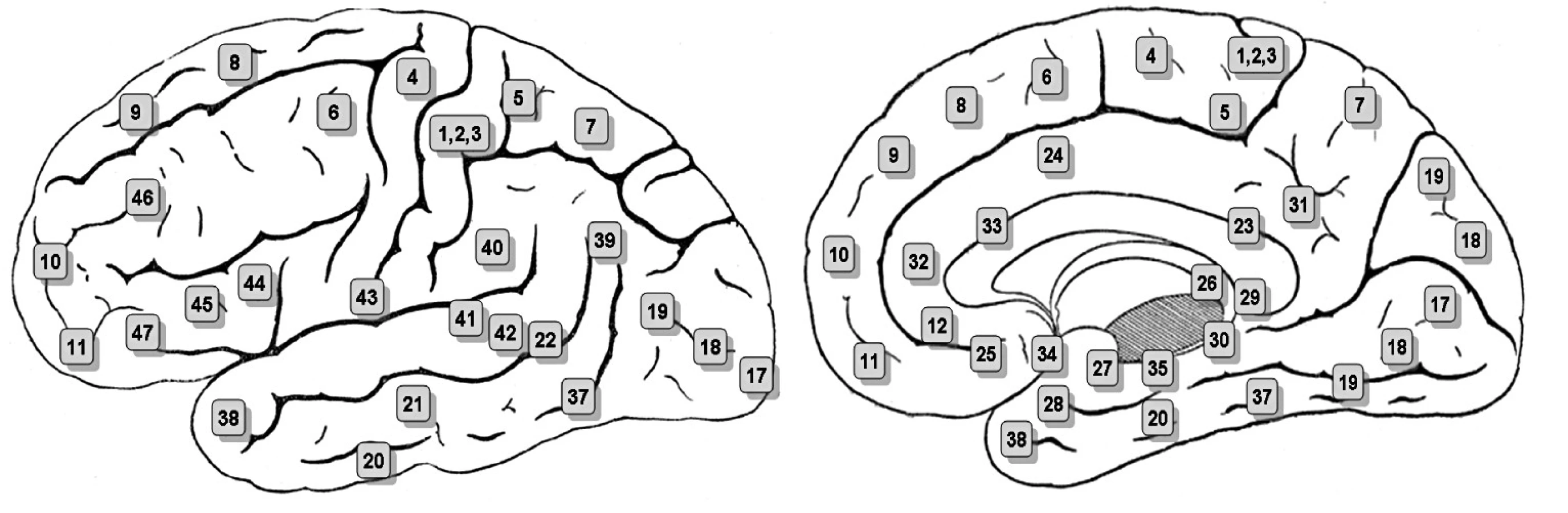
Aktivita mediální orbitofrontální kůry monitoruje, učí se a pamatuje si míru odměny poskytovanou podnětem. Naopak aktivita laterální části téže kůry monitoruje, učí se a pamatuje si míru trestu. Kromě toho existuje předozadní gradient: Složitější podněty, například peněžní zisky nebo ztráty, zpracovává tato část kůry v oblastech bližších frontálnímu pólu, zatímco jednodušší podněty, jako je chuť, zpracovávají kaudálnější („zadní“) části této korové oblasti.
Dalšími korovými oblastmi, které kódují slastné podněty, jsou části střední inzulární (8) a přední cingulární (2) kůry.
Pravostrannou přední inzulární kůru aktivují nabuzující podněty, například bolest. Levostrannou přední inzulární kůru aktivují pozitivní a afiliativní pocity, například pocity matek prohlížejících fotografie vlastních dětí. Větší levostranná než pravostranná aktivace této korové oblasti doprovází mateřskou i romantickou lásku, poslech hlasu vyjadřujícího šťastný pocit, poslech subjektivně příjemné hudby, pohled na usmívající se tvář, vlastní úsměv, pocit radosti, u žen pak je aktivace této korové oblasti úměrná míře prožívání orgasmu. Inzulární kůra byla původně chápána jako oblast viscerální senzorická oblast (8). Inzula a přední cingulární kůra však jsou jak limbické senzorické, tak motorické oblasti, jejichž činnost je podkladem tvorby pocitů. Je pravděpodobné, že se přední inzulární kůra podílí na sebeuvědomování a lze ji považovat za součást implicitního systému lidského mozku (15).
Beckmann et al. (2) užili k mapování konektivity lidské cingulární kůry difuzní traktografii, což je varianta vyšetření magnetickou rezonancí znázorňující průběh systémů vláken bílé hmoty. Dokládá, že na základě zapojení lze cingulární kůru rozdělit do devíti oblastí. Prokazuje propojení cingulární kůry s hipokampem, ventrálním striatem, hypothalamem, parietální, mediální a laterální orbitofrontální, dorzální prefrontální, premotorickou a precentrální kůrou.
Je však nejasné, zda činnost střední a přední části orbitofrontální kůry nebo její části mediální, které slast kódují, je také příčinou pocitu slasti. Poškození této oblasti sice poškozuje rozhodování se vztahem ke slasti, vlastní slastný pocit však mohou mít pacienti s poškozením orbitofrontální kůry uchované, byť je jejich emotivita v tomto směru často nepřiměřená (9, 14).
Hornak et al. (14) vyšetřili emotivitu pacientů po odnětí rozličných částí prefrontální kůry. Předmětem vyšetření byla schopnost identifikovat emoce v hlasu a tváří, sociální chování a subjektivní prožívání emocí. Někteří nemocní s oboustranným, ale i jednostranným poškozením orbitofrontální kůry, případně kombinaci tohoto poškození s lézí přední cingulární kůry a mediální prefrontální kůry v oblasti Brodmanovy area 9 špatně identifikovali emoční výrazy v pozorované tváři a ve slyšeném hlasu, měli poškozené sociální chování a změněné subjektivní prožívání emocí.
Jednostranná nebo oboustranná ohraničená chirurgická léze orbitofrontální kůry tedy poškozuje rozlišování emočního výrazu v pozorované tváři nebo slyšeném hlasu, nemění však významným způsobem sociální chování ani subjektivní prožívání emocí – obojí je změněno při lézi oboustranné. Jednostranné poškození přední cingulární kůry včetně části mediální prefrontální kůry Brodmanovy area 9 poškozuje nejen rozlišování emocí ve tváři a hlasu, znamená i významné změny v subjektivním emočním stavu.
Eudaimonický aspekt
Všechny popsané struktury hédonického mozku, ventrální palidum, ventrální striatum (nc. accumbens) se buď promítají do implicitního neboli defaultního systému lidského mozku, nebo jsou jeho součástí (15). Systém je tvořen některými oblastmi mediální i laterální prefrontální kůry oboustranně, některými částmi kůry cingulární, parietální, korovými oblastmi v rýze mezi horním a středním spánkovým závitem a některými dalšími spánkovými oblastmi. Rozsáhlá literatura dokládá, že jeho činnost je podkladem „niterných podob poznávání“, sebeuvědomování i různých kvalitativně odlišných stavů vědomí, odlišuje se na příklad v průběhu kómatu, vegetativního stavu nebo stavu minimálního vědomí (6, 22).
Lze předpokládat, že implicitní systém může slasti propůjčovat případně individuální smysl. V přední cingulární stejně jako orbitofrontální kůře je vysoká hustota opioidních receptorů. Odchylky stavby, resp. aktivity frontálních součástí implicitního systému například v subgenuální mediální prefrontální kůře a kůře orbitofrontální odpovídají patologickým změnám hédonické zkušenosti u pacientů stižených depresí. Drevets et al. (11) prokazují u pacientů stižených opakovanými epizodami deprese funkční i strukturální odchylky mediální prefrontální kůry, zadní a zevní orbitofrontální kůry, amygdaly, hipokampu a ventromediálních částí bazálních ganglií.
S ohledem na vysokou prevalenci deprese v populaci a odhadovaný celosvětový růst prevalence deprese, která se dostává mezi vedoucí příčiny morbidity a mortality (30) jde o velmi významný výzkumný směr.
Adresa pro korespondenci:
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Thomayerova nemocnice
Vídeňská 800, 140 59 Praha 4 Krč
e-mail: frantisek.koukolik@ftn.cz
Sources
1. Aldridge JW, Berridge KC. Neural coding of pleasure: „rose-tinted glasses“ of the ventral pallidum. In: Kringelbach ML, Berridge KC. (eds.) Pleasures of the brain. New York: Oxford University Press 2010; 62–73.
2. Beckman M, Johansen-Berg H, Rushworth MFS. Connectivity-based parcellation of human cingulate cortex and its relation to functional specialization. J Neurosci 2009; 29 : 1175–1190.
3. Berridge KC, Robinson TE. Parsing reward. Trends Neurosci 2003; 26 : 507–513.
4. Berridge KC, Kringelbach ML. Building a neuroscience of pleasure and well-being. Psychol Well-Being Theory Res Pract 2011; 1(3): 1–26.
5. Blood AJ, Zatorre RJ. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc Natl Acad Sci USA 2001; 98 : 11818–11823.
6. Buckner R, Andrew-Hanna J, Schacter D. The brain´s default network: anatomy, function and relevance to disease. Ann NY Acad Sci 2008; 1124 : 1–38.
7. Carmichael ST, Price JL. Architectonic subdivision of the orbital and medial prefrontal cortex in the macaque monkey. J Comp Neurol 1994; 346 : 366–402.
8. Craig AD. How do you feel – now? The anterior insula and human awareness. Nat Rev Neurosci 2009; 10 : 59–70.
9. Damasio AR. The somatic marker hypothesis and the possible functions of the prefrontal cortex. Phil Trans R Soc B: Biol Sci 1996; 351 : 1413–1420.
10. Damasio A. Hledání Spinozy. Radost, strast a citový mozek. Praha: Dybbuk 2004 (přel. F. Koukolík).
11. Drevets WC, Price JL, Furey ML. Brain structural and functional abnormalities in mood disorders: implications for neurocircuitry models of depression. Brain Struct Funct 2008; 213 : 93–118.
12. Frijda N. On the nature and function of pleasure. In: Kringelbach ML, Berridge KC. (Eds.) Pleasures of the brain. New York: Oxford University Press 2010; 99–112.
13. Georgiadis JR, Kortekaas R, Kuipers R, et al. Regional cerebral blood flow changes associated with clitorally induced orgasms in healthy women. Eur J Neurosci 2006; 24 : 3305–3316.
14. Hornak J, Bramham J, Rolls ET, et al. Changes in emotion after circumscribed surgical lesions of the orbitofrontal and cingulate cortices. Brain 2003; 126 : 1671–1712.
15. Koukolík F. Evoluce a evoluční teorie pro lékaře X. Sebeuvědomování. Prakt. Lék. 2010; 90 : 571–576.
16. Koukolík F. Lidský mozek. Funkční systémy. Norma a poruchy. Třetí, přepracované a doplněné vydání. Praha: Galén 2012; 220–224.
17. Kesebir P, Diener E. In pursuit of happiness: empirical answers to philosophical questions. Perspect Psychol Sci 2008; 3 : 117–125.
18. Kringelbach ML, O´Doherty J, Rolls ET, et al. Activation of the human orbitofrontal cortex to a liquid food stimulus in correlated with subjective plesantness. Cereb Cortex 2003; 13 : 1064–1071.
19. Kringelbach ML. The human orbitofrontal cortex: linking reward to hedonic experience. Nat Rev Neurosci 2005; 6 : 691–702.
20. Kringelbach ML, Berridge KC. Towards a functional neuroanatomy of pleasure and happiness. Trends Cogn Sci 2009; 13 : 479–487.
21. Kringelbach ML. The hedonic brain: a functional neuroanatomy of human pleasure. In: Kringelbach ML, Berridge KC. (Eds.) Pleasures of the brain. New York: Oxford University Press 2010 : 202–221.
22. Laureys S, Owen AM, Schiff ND. Brain function in coma, vegetative state, and related disorders. Lancet Neurol 2004; 3 : 537–546.
23. Miller JM, Vorel SR, Tranguch AJ, et al. Anhedonia after selective bilateral lesion of the globus palidus. Am J Psychiatry 2006; 163 : 786–788.
24. Pessiglione M, Schmidt L, Draganski B, et al. How the brain translates money into force: a neuroimaging study of subliminal motivation. Science 2007; 11 : 904–906.
25. Small DM, Zatorre RJ, Dagher A, et al. Changes in brain activity related to eating chocolate. From pleasure to aversion. Brain 2001; 124 : 1720–1733.
26. Smith KS, Mahler SV, Pecina S, et al. Hedonic hotspots : generating sensory pleasure in the brain. In: Kringelbach ML, Berridge KC. (eds.) Pleasures of the Brain. New York: Oxford University Press 2010 : 27–49.
27. Smith KS, Berridge KC, Aldridge JW. Disentangling pleasure from incentive salience and learning signals in brain reward circuitry. Proc Natl Acad Sci 2011; 108: E255–E264.
28. Steiner JE, Glaser D, Hawillo ME, et al. Comparative expression of hedonic impact: affective reactions to taste by human infants and other primates. Neurosci Biobehav Rev 2001; 25 : 53–74.
29. Völlm BA, de Araujo IE, Cowen PJ, et al. Methamphetamine activates reward circuitry in drug naive human subjects. Neuropsychopharmacology 2004; 29 : 1715–1722.
30. WHO. The world health report (2001). Chap 2,4. Dostupné na: www.who.int/entity/whr/2001/en/whr01_en.pdf
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2013 Issue 1
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
- Metamizole at a Glance and in Practice – Effective Non-Opioid Analgesic for All Ages
Most read in this issue
- Projevy anémií v dutině ústní
- Poruchy metabolismu a nutrice spojené s abúzem alkoholu
- Základní pravidla při provádění pracovnělékařských služeb ve světle zákona o specifických zdravotních službách
- Základy kognitivní, afektivní a sociální neurovědy XXV. O štěstí – hédonický mozek