
Mastné kyseliny 2. Fyziologický a klinický význam
Fatty acids – 2. Clinical and physiological significance
Fatty acids play multiple roles in humans and other organisms. In triglycerides they are the source of metabolic energy, in adipose tissue they serve also as temperature and mechanical isolators, in the form of phospholipids they are structural components of membranes. Fatty acids originating from the sn-2 glycerol carbon of phosphatidylcholine can influence the activity of diglycerides as second messengers. Unsaturated FA with 18-20 carbon atoms are precursors of prostaglandins, leucotrienes and thromboxanes, which have a broad scale of regulatory properties and have autocrine as well as paracrine effects. Fatty acids are ligands of several nuclear receptors, which take part in the subcellular control of a number of metabolic pathways. Covalent modification of proteins by FA (acylation) enables FA incorporation into the membranes. Number of pathological stages is accompanied with changes in fatty acid composition, often expressed as decreased content of unsaturated and increased content of saturated fatty acids (e.g. dyslipidemia, malnutrition, inflammation and inherited diseases). Polyunsaturated fatty acids as dietary supplements are used in prevention and in the therapy of cardiovascular diseases and other metabolic disturbances.
Key words:
fatty acids, membranes, eicosanoids, metabolism.
Autoři:
E. Tvrzická; B. Staňková; M. Vecka; A. Žák
Působiště autorů:
Univerzita Karlova v Praze, 1. lékařská fakulta, IV. interní klinika VFN
Vyšlo v časopise:
Čas. Lék. čes. 2009; 148: 116-123
Kategorie:
Přehledový článek
Souhrn
Mastné kyseliny mají v organismu řadu důležitých úloh. V triglyceridech jsou zdrojem metabolické energie, v tukové tkáni slouží jako mechanické a tepelné izolátory, ve formě fosfolipidů jsou strukturními složkami membrán. Mastná kyselina pocházející z prostředního uhlíkového atomu glycerolu fosfatidylcholinu může ovlivnit aktivitu diglyceridu jako druhého posla. Nenasycené mastné kyseliny s 20 atomy uhlíku jsou prekurzory eikosanoidů, které mají široké pole autokrinních i parakrinních účinků. Mastné kyseliny jsou ligandy některých nukleárních receptorů, které se účastní mnoha metabolických procesů. Kovalentní modifikace proteinů (acylace) umožňuje jejich inkorporaci do membrán. Řada patologických stavů je doprovázena změnami ve složení mastných kyselin, velmi častý je pokles obsahu nenasycených a nárůst obsahu nasycených mastných kyselin (např. u dyslipidémie, malnutrice, zánětu, vrozených chorob). Vícenenasycené mastné kyseliny jsou ve formě dietních doplňků využívány jak k prevenci, tak i k léčbě kardiovaskulárních onemocnění a dalších metabolických poruch.
Klíčová slova:
mastné kyseliny, membrány, eikosanoidy, metabolismus.
Úvod
Mastné kyseliny (FA) mají v živých organismech řadu úloh. Jako součást fosfolipidů (PL) tvoří základní strukturu všech buněčných membrán, v triglyceridech (TG) představují významný zdroj energie; v tukové tkáni slouží jako zásoba energie i tepelná izolace. Mohou být takto skladovány prakticky v neomezeném množství, což můžeme pozorovat například u obézních jedinců a některých savců připravujících se na zimní spánek.
Nenasycené FA s 18–20 atomy uhlíku jsou prekurzory prostaglandinů, leukotrienů a tromboxanů, které mají široký rozsah regulačních vlastností, s autokrinními i parakrinními účinky. Mastné kyseliny jsou rovněž ligandy několika nukleárních receptorů, které kontrolují řadu metabolických pochodů na subcelulární úrovni. Kovalentní modifikace proteinů mastnými kyselinami umožňuje jejich inkorporaci do membrán. Mastné kyseliny s 20 a 22 uhlíkovými atomy jsou prekurzory dalších autakoidů: dokosanoidů, resolvinů, lipoxinů a neuroprotektinů (1). Hydroxy FA jsou aktivátory některých nukleárních faktorů (např. NF-κB, AP-1 and TNF-α) a jsou zodpovědné za expresi prozánětlivých cytokinů (např. IL-1, IL-6, IL-8 a TNF-α) a adhezních molekul (např. ICAM-1, VCAM-1 a ELAM-1).
Fyziologická úloha mastných kyselin
Mastné kyseliny plní v lidském organismu několik důležitých funkcí.
Zdroj energie
Mastné kyseliny ve formě TG jsou základním zdrojem metabolické energie. Oproti proteinům a sacharidům mají více než dvojnásobnou kalorickou hodnotu a k jejich skladování je třeba menšího množství vody. Energetická hodnota FA s dlouhým a velmi dlouhým řetězcem (LCFA a VLCFA) je 9,1 kcal/ g, pro FA se středním řetězcem (MCFA) to je 7,0 kcal/ g; FA s krátkým řetězcem (SCFA) představují energetický zdroj enterocytů a kolonocytů. Esenciální FA v molekulách TG jsou využívány pro další metabolické procesy, např. pro tvorbu signálních molekul (2). Hydrofobní charakter TG je navíc užitečný pro transport i skladování důležitých lipofilních molekul (vitaminy A, D, E a K).
Izolátory
Tkáně s vysokým obsahem TG (subkutánní a viscerální tuk) slouží také jako tepelné a mechanické izolátory. Navíc VLCFA jsou strukturálními složkami ceramidů, které snižují prostupnost kůže pro vodu a mají nezastupitelné místo pro udržení vodní bilance (2). Zvláštním případem je přítomnost VLCFA v sekretu Meibomovy žlázy, kde tvoří bariéru mezi slzným filmem a kožními lipidy, a brání tak nadměrnému odpařování slz.
Nervová tkáň obsahuje řadu izolačních lipidů, které obklopují axony, a umožňují tak rychlý přenos informace. Čím silnější je myelinová vrstva, tím rychleji postupuje signál axonem.
Chemická struktura buněčných membrán
Mastné kyseliny ve formě PL jsou strukturními složkami všech buněčných membrán. Jejich složení ovlivňuje sílu i prostupnost membrán, a tím i aktivitu membránově vázaných proteinů (enzymů, iontových kanálů, receptorů a transportérů). Molekula PL obsahuje hydrofilní „hlavičku“ (cholin, etanolamin, serin, inositol), spojenou ve většině případů prostřednictvím kyseliny fosforečné s hydrofobním koncem (1 nebo 2 molekuly FA). Tato vlastnost umožňuje uspořádání tzv. lipidové dvojvrstvy, která má uvnitř hydrofobní část molekul a na vnější straně, orientované k vodnému prostředí, hydrofilní hlavičku. Pevnost membrány je zvýšena přítomností molekul cholesterolu a specifických proteinů. Čím vyšší je jejich obsah, tím nižší je propustnost (fluidita) membrány. Dalším faktorem, který fluiditu membrány ovlivňuje, je počet dvojných vazeb v molekulách FA. Ty jsou v membránách přítomny v převážné většině v cis-konfiguraci, každá vazba tedy způsobuje odklon řetězce od přímého směru pod úhlem cca 60° (dvojná vazba svírá úhel 120°). Řetězce tak zaujímají větší prostor, což se projeví zvýšenou fluiditou. Naopak přítomnost nasycených FA či nenasycených s trans konfigurací dvojných vazeb vnese do membrány rovné řetězce, které zaujímají menší prostor a fluidita membrány se sníží. Zvýšení počtu dvojných vazeb při stejném obsahu PL ale ovlivňuje propustnost membrány v menší míře než zvýšení obsahu PL vůči ostatním složkám (2).
Určitou obdobou buněčné membrány je obal lipoproteinu. Obsahuje rovněž PL, cholesterol a specifické bílkoviny, zvané apolipoproteiny. Molekuly PL jsou orientovány polární hlavičkou vně částice, a umožňují tak rozpustnost lipoproteinů ve vodném prostředí krevní plazmy. Hydrofobní řetězce FA jsou orientovány dovnitř částice, jejíž jádro je tvořeno estery cholesterolu a TG. Propustnost „membrány“ lipoproteinů stoupá v řadě HDL < LDL < VLDL (3).
Chemická struktura druhých poslů
Informace v živém organismu je přijímána ve formě fyzikálních nebo chemických signálů (2). Vnitřní chemické signály, mezi které patří především hormony, nervové mediátory a růstové faktory, vyvolávají po vazbě na specifické receptory aktivaci látky zvané druhý posel, což může být např. cyklický adenosinmonofosfát (cAMP), cyklický guanosinmonofosfát (cGMP), ionty kalcia, dále pak diglycerid (DG), inositoltrifosfát nebo oxid dusnatý. Například acetylcholin (signální molekula) ve vazbě na membránový receptor aktivuje fosfolipasu C, ta hydrolýzou fosfatidylinositol 4,5-bisfosfátu (PIP2) v membráně vytvoří fosfatidylinositol 1,4,5-trisfosfát (PIP3) a DG (oba působí jako druzí poslové); PIP3 uvolňuje ionty kalcia ze sarkoplazmatického retikula, DG aktivuje proteinkinázu C. Aktivitu DG jako druhého posla může ovlivnit FA pocházející z prostředního (sn-2) uhlíkového atomu glycerolu fosfatidylcholinu, jak schematicky ukazuje obrázek 1.
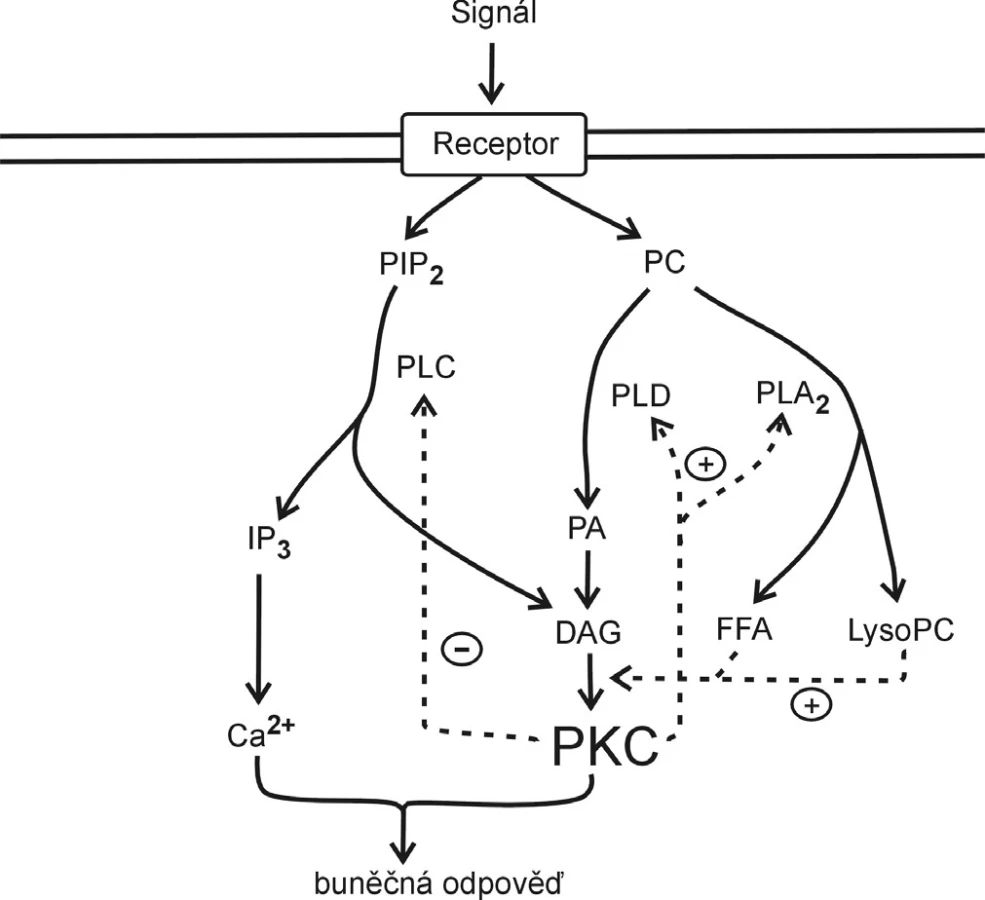
Prekurzory eikosanoidů a substrát pro lipoperoxidaci
Eikosanoidy jsou základní hormony či regulační molekuly, které jsou přítomny u většiny forem života. Na rozdíl od endokrinních hormonů, které cestují krevním řečištěm, jsou eikosanoidy autokrinní nebo parakrinní faktory. Ovlivňují jak aktivity buněk, ve kterých jsou syntetizovány, tak i přilehlých buněk vazbou na 7TM receptory. Název eikosanoidy pochází z řeckého „eikosi“ = dvacet, protože obsahují 20 uhlíkových atomů v molekule. Nejznámějšími eikosanoidy (nejvíce prostudovanými) jsou prostaglandiny (PG), tromboxany (TX) a leukotrieny (LT) (4).
Prostaglandin- a tromboxan-syntetázy, enzymy, které katalyzují první krok metabolického procesu vedoucího přes endoperoxidy PGG2 a PGH2 k prostacyklinu, PG a TX, sestávají z cyklooxygenázy (COX) a hydroperoxidázy. Typickým inhibitorem COX je kyselina acetylosalicylová (Aspirin), protizánětlivý i antitrombotický lék, stejně jako další nesteroidní protizánětlivé léky(NSAID); tyto léky ireverzibilně blokují syntézu eikosanoidů, a ovlivňují tak řadu signálních funkcí. Výsledkem tohoto kroku je potlačení zánětu, horečky, bolesti a srážení krve.
Prostaglandiny stimulují zánětlivý proces, regulují tok krve k jednotlivým orgánům, kontrolují transport iontů membránami, modulují synaptické transmise a indukují spánek.
Tromboxany jsou vazokonstriktory a potenciální hypertenzivní faktory, které též usnadňují agregaci krevních destiček. Podle tohoto srážlivého účinku jsou i pojmenovány (trombóza – trombocyty). Tromboxan A2, produkovaný aktivovanými destičkami, má protrombotické vlastnosti (stimuluje aktivaci nových destiček i jejich agregaci). Index u zkratky PG a TX představuje počet dvojných vazeb v molekule, zbývající dvojné vazby prekurzoru jsou využity pro tvorbu cyklo- nebo oxy-derivátů.
Endogenním eikosanoidem je anandamid (arachidonyl ethanolamid), který se specificky váže na kanabinoidní receptory (CB); CB1 jsou exprimovány v mozku, gastrointestinálním traktu a tukové tkáni, CB2 jsou exprimovány v imunitním systému (5).
Lipoxygenáza (LOX) katalyzuje počáteční krok metabolického procesu vedoucího k LT. Název leukotrien reflektuje skutečnost, že byly poprvé objeveny v leukocytech, a že obsahují 3 konjugované dvojné vazby (4).
Prekurzory eikosanoidů jsou vícenenasycené FA (PUFA) odštěpené z sn-2 atomu uhlíku glycerolu fosfolipidů, jmenovitě arachidonová (AA), eikosapentaenová (EPA) a dihomo-γ-linolenová (DHGLA). Schéma metabolismu AA ukazuje obrázek 2. Mastné kyseliny jsou odštěpeny působením fosfolipasy A2 a transformovány působením COX typu 1 a 2 na PG a TX, a pomocí LOX typu 5, 8, 12 a 15 na LT a kyseliny hydroxyeikosatetraenovou (HETE), hydroperoxyeikosatetraenovou (HPETE) a hydroxyeikosapentaenovou (HEPE). Některé trihydroxy deriváty EPA se nazývají lipoxiny (LX) nebo resolviny (Rv). Jako substráty pro lipoperoxidaci však slouží také kyseliny linolová (18:2n-6), eikosadienová (20:2n-6), Meadova (20:3n-9) a dokosahexaenová (DHA).
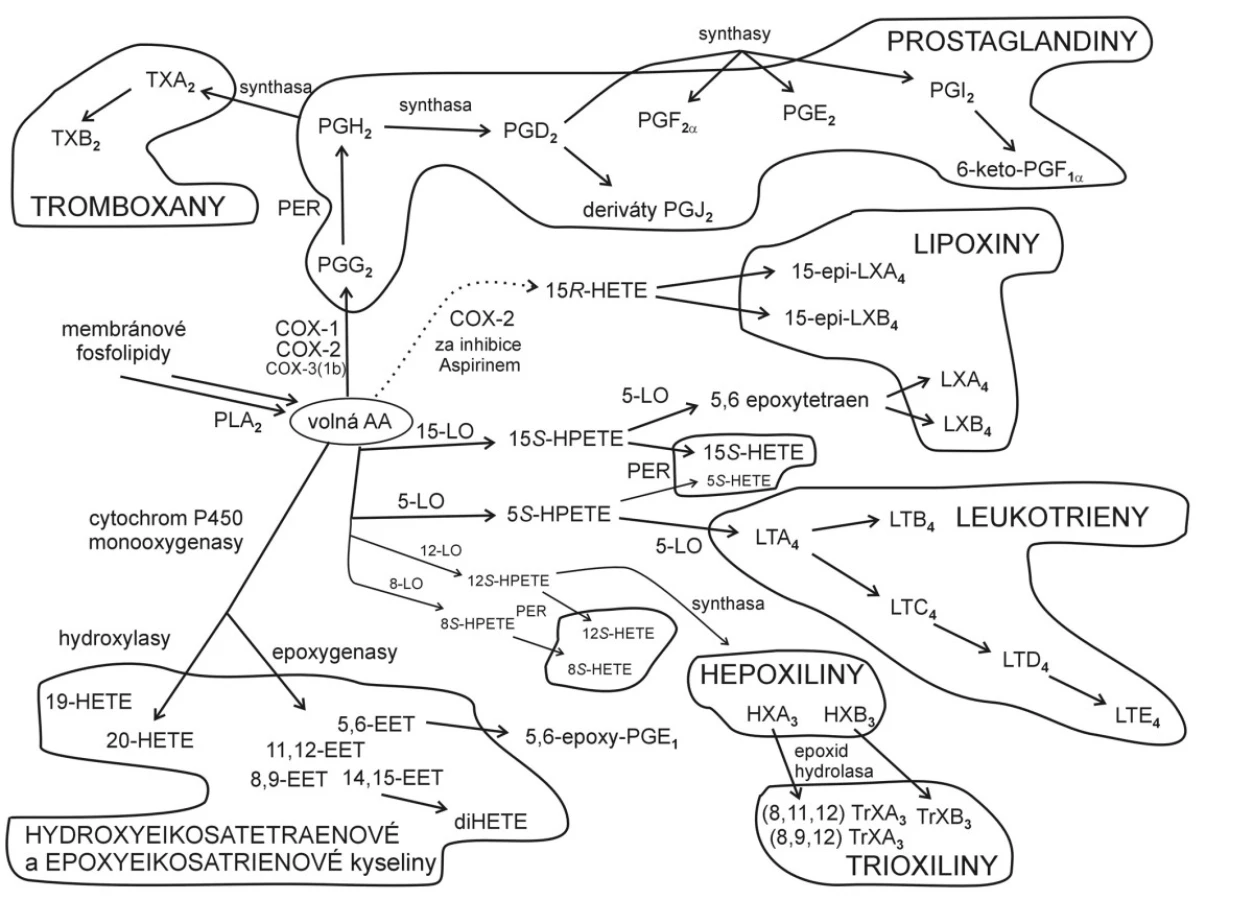
Kyselina arachidonová je prekurzorem PG série 2 a LT série 4, EPA je prekurzorem PG série 3 a LT série 5, DHGLA je prekurzorem PG série 1 a LT série 3. Prostaglandiny a LT odvozené od AA jsou často syntetizovány jako odpověď na určitý akutní stav (úraz, stres), zatímco PG a LT odvozené z EPA modulují účinky PG odvozených od AA (obvykle ve stejné cílové buňce). Prostaglandiny odvozené z EPA se tvoří pomaleji a jejich úkolem je tlumit účinky zvýšených hladin PG odvozených z AA; jejich přiměřená produkce pravděpodobně chrání před srdečním infarktem, mozkovou příhodou, a dále před některými zánětlivými onemocněními (artritida, lupus, astma). Eikosapentaenová kyselina prostřednictvím svých mono- a trihydroxy derivátů snižuje produkci prozánětlivých cytokinů (např. IL-1β, TNF α). Potlačení produkce eikosanoidů odvozených z PUFA n-6 pomocí PUFA n-3 je pravděpodobně způsobeno kompeticí obou řad PUFA o společné enzymy, které se účastní elongace a desaturace kyseliny linolové (LA) na AA.
Specifické markery lipoperoxidace LA jsou kyseliny 9 a 13-hydroxy oktadekadienová (9-HODE a 13-HODE), kyselina 13-keto-oktadeka-9,11-dienová (13-OXO) a kyselina 13-hydroperoxy oktadekadienová (13-HPODE), které jsou významnými složkami oxidovaných LDL a účinnějšími aktivátory NF-κB než 13-HODE. Dalšími významnými markery lipoperoxidace jsou malondialdehyd, 4-hydroxy-nonenal a F2-izoprostany.
Produkty neenzymatické oxidace AA a dalších dvacetiuhlíkatých FA (izoprostany) jsou sledovány jako zástupné (pomocné) markery oxidativního stresu. Cyklické sloučeniny odvozené z kyseliny α-linolenové (ALA), EPA a DHA se nazývají neuroprostany. Je známa rovněž tvorba necyklických produktů nazývaných izoketaly (6).
Spontánní reakce FA s molekulárním kyslíkem jsou předmětem studia vzhledem k účasti reakčních produktů v destruktivních biologických pochodech. Autooxidační procesy mají za následek tvorbu řady sloučenin, jako jsou hydroperoxidy, FA se 2 a více dvojnými vazbami tvoří cyklické produkty. Tyto sloučeniny nejsou stálé a prostřednictvím hydroxy derivátů tvoří stabilní sloučeniny s karbonylovou skupinou. Některé z nich slouží jako markery oxidačního stresu (7).
Acylace proteinů
Acylace proteinů je důležitým způsobem kovalentní modifikace proteinů. Umožňuje jejich asociaci s biologickými membránami, a tím zvýšení jejich strukturální stability, dále pak interakce protein/protein i katalytickou aktivitu (8). Modifikované proteiny plní v organismu řadu funkcí.
V acylaci proteinů se uplatňují dva základní procesy – modifikace kyselinou myristovou, která byla již detailně prostudována, a kyselinou palmitovou, jejíž mechanismy zatím nejsou přesně známy.
Myristoylace probíhá na části proteinu zakončeném sekvencí methionin-glycin. Koncový methionin je odstraněn kotranslačně methionin amino-peptidázou a myristát je spojen s glycinem amidovou vazbou. Reakce je ireverzibilní a probíhá za katalýzy N-myristoyl transferázou.
Palmitoylace je realizována posttranslační reverzibilní thioesterifikací cysteinu, která není přímo spojena se vznikem proteinu, a je využívána i buňkami bez transkripčních aktivit (např. destičky). Protein může být deacylován působením enzymu palmitoyl protein thioesterázy (8).
Suplementace PUFA n-3 ovlivňuje příznivě acylaci proteinů, funkci iontových kanálů v myokardu a elektrickou stabilitu. Snižuje riziko náhlé kardiální smrti i fatálního infarktu myokardu (DART Study, Lyon Heart Study and GISSI Prevenzione Trial) (9–11).
Signální funkce a modulace genové transkripce
Buněčná signalizace zajišťuje přenos informace mezi buňkami jednotlivých soustav a její narušení má zásadní úlohu v patogenezi onemocnění. Signální dráhy mohou být ovlivněny některými geny, nukleárními faktory či receptory. Mastné kyseliny jako ligandy těchto receptorů či modulátory genové transkripce mohou buněčnou signalizaci významně ovlivnit (12).
Vliv FA (zvláště kyseliny stearové) na uvolňování inzulínu z β-buněk Langerhansových ostrůvků je jednak aktivační (akutní expozice FA většinou podporuje uvolňování inzulínu), jednak inhibiční (trvale zvýšené hladiny FA zhoršují inzulínovou rezistenci). Uvolňování inzulínu je podporováno přednostně nasycenými FA (SFA), na rozdíl od PUFA. Mechanismus odpovědi β-buněk není znám, uvažuje se o dosud neznámém metabolitu (receptoru) FA, který vzniká působením inzulínu na protein vázající sterolový regulační element (SREBP-1c) (13).
Nenasycené FA s dlouhým řetězcem (C20-22) a jejich metabolity jsou rovněž ligandy nukleárních receptorů některých transkripčních faktorů:
- receptory aktivované peroxizomovými proliferátory (PPAR-α, PPAR-β/δ, PPAR-γ1 a PPAR-γ2),
- jaterních X receptorů (liver X receptors, LXR) typu α a β,
- jaterních nukleárních faktorů (HNF) 4α,
- SREBP-1 a 2.
Neesterifikované LCPUFA inhibují (jako ligandy) PPAR a LXR. Zpětná regulace HNF-4 je realizována thioestery acyl-CoA PUFA. Naproti tomu SREBP, které jsou senzory intracelulárních konverzí lipidů, se účastní syntézy cholesterolu (SREBP-1a SREBP-2), FA a TG (SREBP-1c).
Mechanismus působení PPAR ukazuje obrázek 3. Transkripční faktory interagují s odpovídajícími vazebnými sekvencemi DNA, které kódují geny metabolicky významných bílkovin (enzymů, aktivátorů či inhibitorů lipidové a glukózové homeostázy, karcinogeneze, homeostázy tukové tkáně a dalších) (14).
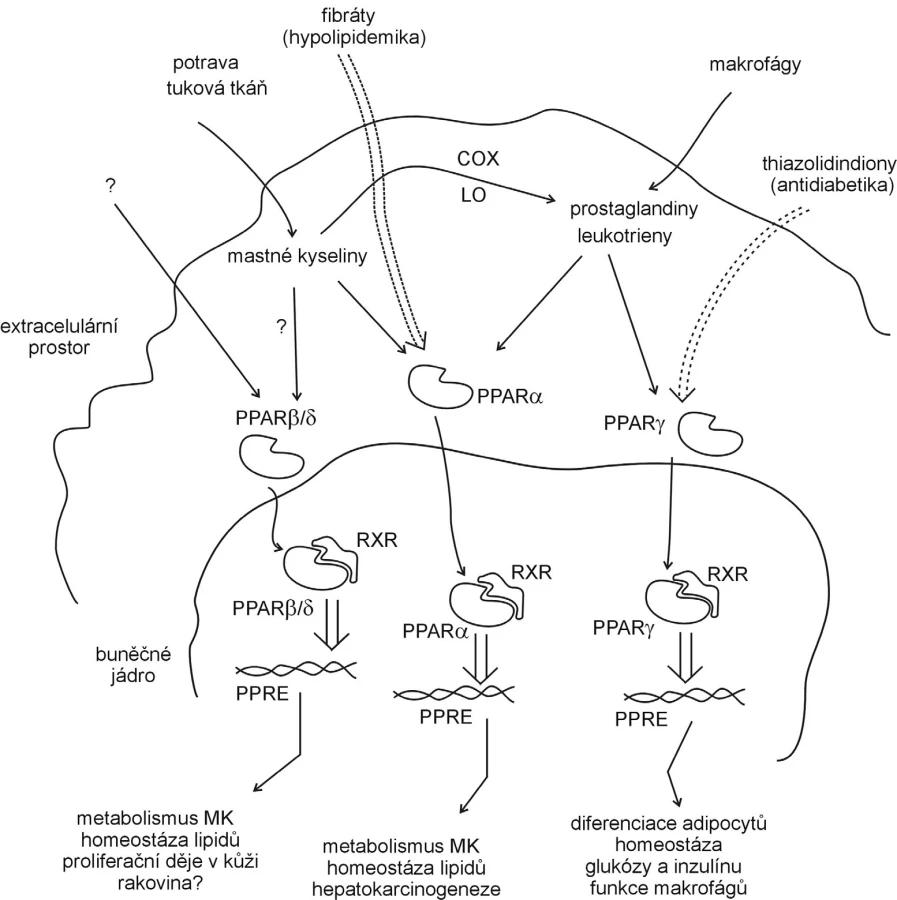
Ligandy receptorů
V polovině minulého století se začaly objevovat studie amidových derivátů FA. U N-palmitoyletanolaminu ze sóji a vaječného žloutku bylo prokázáno působení jako protizánětlivého faktoru. Etanolamin, amonium a některé bioaktivní aminy jsou donory aminoskupiny při reakci s FA. Výsledné amidy reagují jak s kanabinoidními, tak s vaniloidními receptory. Mozkové kanabinoidní receptory se účastní signálních procesů paměťových, pohybových, emocionálních a nociceptivních. Naopak periferní kanabinoidní receptory se účastní mimo jiné modulace imunokompetentních buněk (15).
Etylestery FA, produkty neoxidativního metabolismu etanolu, jsou produkovány specifickými syntázami. Koncentrace etylesterů FA je specifickým markerem chronického abúzu alkoholu. Tyto estery mohou být ligandy PPAR, transkripčních faktorů κB a aktivačního proteinu 1 (16).
Receptory pro faktor aktivující destičky (PAF, 1-O-alkyl-2-acetyl-sn-glycerol-3-fosfocholin) jsou vysoce specifické pro molekulu PAF, která obsahuje v sn-1 poloze glycerofosfocholinu éterovou vazbu a v sn-2 poloze zbytek krátké FA (17).
Nereceptorové interakce mezi proteiny a mastnými kyselinami
Některé FA (olejová, palmitolejová, arachidonová) jsou účinnými odpřahujícími prostředky buněčné komunikace na úrovni mezibuněčných iontových kanálů. Mechanismus působení spočívá ve změně interakce mezi lipidy a spojujícími proteiny buněk, zvláště myocytů (18).
Mastné kyseliny ovlivňují i iontové kanály; například kalciové kanály ventrikulárních myocytů jsou přímo aktivovány LC PUFA. Byl popsán vliv DHA na aktivitu draslíkového kanálu usměrňující tok proudu intracelulárně jako důsledku jejího účinku na vedlejší podjednotku kalciového kanálu. Sodné kanály vykazují rozdíly mezi srdeční a svalovou izoformou v závislosti na typu inhibující FA. Těmito interakcemi jsou pravděpodobně zapříčiněny antiarytmické účinky n 3 PUFA (19).
Patofyziologie mastných kyselin
Za fyziologických podmínek je profil FA v lipoproteinech i buňkách jednotlivých tkání určen především geneticky (živočišným druhem a typem tkáně), je však ovlivněn i dietním příjmem (FA i dalších složek diety), působením hormonů a dalšími faktory (teplota, radiace, viry). Zmíněné faktory ovlivňují aktivity elongáz a desaturáz FA, které mají určující vliv na jejich konečné složení (20). Přehled působení těchto faktorů na jednotlivé enzymy metabolismu FA udává tabulka 1.
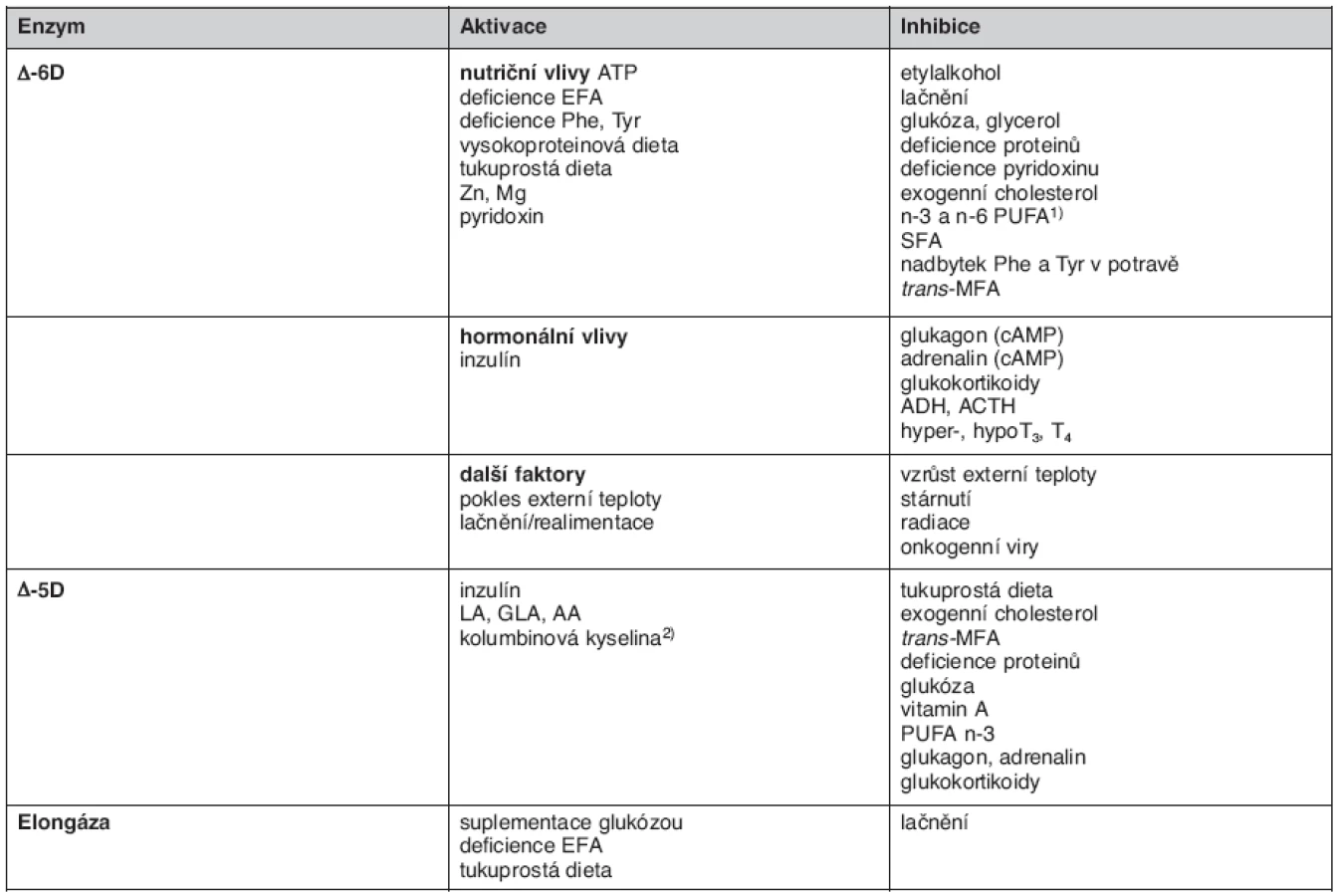
Řada patologických stavů je spojena s charakteristickými změnami ve složení FA, které se projeví většinou zvýšeným obsahem SFA a níženým obsahem PUFA. Takto se projevuje například dyslipidémie, karenční stavy a malnutrice, zánět, oxidativní stres, poruchy metabolismu (metabolický syndrom, diabetes mellitus 2. typu aj.). Není bez zajímavosti, že metabolické poruchy s tak odlišným projevem, jako je extremní hubenost při mentální anorexii oproti obezitě u metabolického syndromu, mají obdobné změny ve složení FA. Charakteristickým znakem obou poruch je vedle zvýšeného obsahu SFA i zvýšený obsah kyseliny palmitolejové (která je markerem liponeogeneze) jako důsledek zvýšené aktivity ∆9 desaturázy kyseliny palmitové a snížený obsah kyseliny linolové, který je pravděpodobně výslednicí několika faktorů (snížený příjem, zvýšená peroxidace a ß-oxidace, zvýšená přeměna na AA pro syntézu eikosanoidů) (21, 22).
Nedostatek esenciálních FA je znám od 30. let minulého století (23) a v experimentu na zvířatech byl spojován s retardací růstu a zvýšenými transepidermálními ztrátami vody v důsledku vzestupu permeability kůže. U samců i samic se karence EFA projevovala sterilitou. Snížil se obsah kyseliny arachidonové, která je obsažena především ve fosfolipidech buněčných membrán a obalu lipoproteinů, a je mj. prekurzorem eikosanoidů (24–26). Organismus v této situaci zvýší desaturaci a elongaci kyseliny olejové na tzv. Meadovu kyselinu (20:3n-9) ve snaze zachovat fluiditu membrán a získat alternativní prekurzor pro syntézu eikosanoidů (27). S poruchou mikrotokových vlastností buněčných membrán, resp. poruchou stability biomembrán souvisí zřejmě popisovaná zvýšená fragilita kapilár a hematurie. Experimentální zvířata vykazují dále zvýšenou spotřebu potravy s negativní dusíkovou bilancí, pokles produkce ATP parenchymatózních orgánů (játra, myokard), který se projevuje sníženou kontraktilitou myokardu a abnormálními zálomy komplexu QRS na EKG. Deficit EFA v játrech vede k poruše transportu cholesterolu se sekundární dyslipidémií a zpomalením reverzního transportu cholesterolu. Somatické příznaky doplňuje porucha adaptace na tmu (dysopsie), snížená zraková ostrost i senzorické a motorické neuropatie (28–30). V 90. letech byly definovány patofyziologicky a klinicky rozdíly karence PUFA n-3 a PUFA n-6, které jsou uvedeny v tabulce 2.

V souvislosti s nevhodným složením dietního tuku je zmiňován i obsah trans FA. Tyto kyseliny jsou považovány za více aterogenní než SFA. Jejich hlavním představitelem je kyselina transvakcenová, která vzniká bakteriálním působením v zažívacím traktu přežvýkavců a je obsažena v mléce a mléčných produktech. Lidský organismus je pravděpodobně na „dlouhodobý“ přísun této kyseliny v mléčných výrobcích za historicky dlouhé období již adaptován. Metabolit kyseliny transvakcenové – kyselina rumenová – je zmíněn v 1. části této práce (31).
Dalším představitelem trans FA je kyselina elaidová, která vzniká díky nekvalitnímu katalyzátoru při ztužování rostlinných olejů na pokrmové tuky. Díky vyspělým technologiím i požadavku deklarace obsahu trans FA ve výrobku se jejich obsah postupně minimalizuje.
Terapeutické využití vícenenasycených mastných kyselin
Dietní suplementace PUFA n-3 bývá spojována s obavami z oxidativního stresu a zvýšené oxidativní modifikace LDL ve srovnání s PUFA n-6. Je však třeba vzít v úvahu i nové názory o inkorporaci PUFA n-3 do fosfolipidů, která vede ke konformačním změnám a nižší náchylnosti dvojných vazeb k lipoperoxidaci než u PUFA n-6. Peroxylové radikály odvozené z EPA jsou více hydrofilní než radikály vzniklé z LA; ochotněji difundují obalem lipoproteinů a na povrchu lipoproteinu je radikálová reakce rychleji ukončena (32, 33).
Zvýšený dietní příjem PUFA n-3 má, na rozdíl od PUFA n-6, za následek zvýšenou transkripci antioxidačních enzymů – odpřahujícího proteinu 2, glutathion transferázy 2τ, superoxid dismutázy, a potlačenou transkripci enzymů, které se účastní tvorby reaktivních kyslíkatých a dusíkatých sloučenin (RONS). Suplementace PUFA n-6 zvyšuje aktivitu glutathion peroxidázy, superoxid dismutázy a katalázy (33, 34).
V současné době mají některé země zaregistrovány etyl estery PUFA n-3 jako léčivo pro dyslipidémie (např. USA, Rakousko). Je třeba zdůraznit, že dávka PUFA n-3 potřebná pro indukci genové exprese je 3–5× vyšší než dávka pro prevenci deficience EFA. Minimální denní dávka pro prevenci zahrnuje 400 mg ALA a 200 mg EPA i DHA. Doporučené dávky pro LA se pohybují mezi 5 a 14 g/den, pro ALA 1,5–2 g/den, a pro sumu EPA a DHA 0,3 až 1,6 g/den (35).
Enterální suplementace lipidovou emulzí obsahující PUFA n-3 a triglyceridy o střední délce řetězce (MCT) má příznivé účinky u pacientů s chylomikronemickým syndromem kombinovaným s těžkou akutní pankreatitidou. Vícenenasycené FA jsou také využívány pro kombinovanou terapii s fibráty, statiny či kyselinou nikotinovou v případech, kdy monoterapie není dostatečně účinná (36).
Užívání PUFA n-3 je výhodné i pro prevenci koronární srdeční choroby. Využívá se jejich antiarytmogenních a antitrombotických účinků ve spojení s normalizací endoteliální dysfunkce a profilu krevních lipidů. Doporučená denní dávka PUFA n-3 v sekundární prevenci je 1 g EPA a DHA (podle American Heart Association). Při léčbě metabolického syndromu mají PUFA n-3 příznivé účinky na jeho kardiovaskulární a hemodynamickou složku, i když neovlivňují inzulínovou rezistenci. Nezanedbatelné jsou v tomto směru i jejich mírné hypotenzivní a antitrombotické účinky (28).
Izolované etylestery DHA mohou zlepšit klinické i biochemické parametry u dětí s vrozenými poruchami biogenze peroxizomů, jako je Zellwegerův syndrom, novorozenecká adrenoleukodystrofie či infantilní forma Refsumovy choroby (33, 37).
Enterální výživa i parenterální výživa obsahující EPA má široké použití u kriticky nemocných; příznivé účinky byly popsány u syndromu šokové plíce (ARDS), diseminované intravaskulární koagulace a dalších stavů (jaterní steatóza, biliární sludge, cholelitiáza). U pacientů po elektivních chirurgických výkonech na zažívacím ústrojí pro karcinom snižuje tato výživa gastrointestinální a infekční komplikace až o 50 % (35, 36, 38, 39).
Ztráty kosterního svalstva u některých typů proteino-energetické malnutrice jsou spojeny se zvýšeným katabolismem proteinů vlivem TNF-α, glukokortikoidů a faktoru indukujícího proteolýzu (PIF). Suplementace EPA zpomaluje degradaci kosterních svalů pravděpodobně inhibicí PIF. V experimentálních studiích byl popsán inhibiční účinek EPA na proteazómovou aktivitu (40, 41).
Závěr
Mastné kyseliny jsou důležitou součástí všech organismů rostlinné i živočišné říše. Jejich složení je na všech stupních dáno především geneticky, jako významná složka potravního řetězce však mohou toto složení částečně ovlivnit. O mastných kyselinách a jejich působení v organismu máme již velmi mnoho poznatků, přesto ty nejvýznamnější, jako je např. tvorba různých eikosanoidů a dalších působků (dokosanoidy, resolviny) a jejich působení, modulace genové transkripce, souvislost s expresí genů a jejich polymorfismy stále čekají na detailní výzkum. Úspěšnost je do značné míry závislá na vývoji specializovaných metodik, které tyto poznatky umožní.
Zkratky
AA – kyselina arachidonová (arachidonic acid)
ALA – kyselina α-linolenová (α-linolenic acid)
AP – aktivátorový protein
ARDS – syndrom dechové tísně dospělých (adult respiratory distress syndrome)
ATP – adenosintrifosfát
cAMP – cyklický adenosinmonofosfát
CB – kanabinoidní receptor
cGMP – cyklický guanosinmonofosfát
COX – cyklooxygenáza
DART – Diet and Reinfarction Trial
DG – diglyceridy
DHA – kyselina dokosahdexaenová (docosahexaenoic acid)
DHGLA – kyselina dihomo-γ-linolenová (dihomo-γ-linolenic acid)
EFA – esenciální mastné kyseliny (essential fatty acids)
ELAM – endoteliální leukocytová adhezní molekula
EPA – kyselina eikosapentaenová (eicosapentaenoic acid)
FA – mastné kyseliny (fatty acids)
GISSI – Gruppo italiano per lo Studio della Sopravvivenza nell’Infarto Miocardico
HDL-C – cholesterol v lipoproteinu o vysoké hustotě (high density lipoprotein cholesterol)
HETE – hydroxyeikosatetraenová kyselina
HEPE – hydroxyeikosapentaenová kyselina
HODE – hydroxyoktadekadienová kyselina
HNF – jaterní nuklerání faktor (hepatic nuclear factor)
HPETE – hydroperoxyeikosatetraenová kyselina
HPODE – hydroperoxyoktadekadienová kyselina
ICAM – intercelulární adhesní molekula
IL – interleukin
LA – kyselina linolová (linoleic acid)
LCFA – mastné kyseliny s dlouhým řetězcem (long chain fatty acids)
LDL-C – cholesterol v lipoproteinu o nízké hustotě (low density lipoprotein cholesterol)
LOX – lipoxygenáza
LT – leukotrieny
LX – lipoxiny
LXR – jaterní receptor X (liver X receptor)
MCFA – mastné kyseliny se středním řetězcem (medium chain fatty acids)
MCT – triglyceridy se středním řetězcem (medium chain triglycerides)
MFA – monoenové mastné kyseliny
NFκB – nukleární faktor κB
NSAID – nesteroidní protizánětlivé léky (nonsteroidal antiinflammatory drugs)
PAF – faktor aktivující destičky (platelet activating factor)
PG – prostaglandiny
PIF – faktor indukující proteolýzu (proteolysis inducing factor)
PIP2 – fosfatidylinositol 4,5-bisfosfát
PIP3 – fosfatidylinositol 1,4,5-trisfosfát
PL – fosfolipidy
PPAR – receptory aktivované peroxizomovými proliferátory (peroxisome proliferator activated receptor)
PUFA – vícenenasycené mastné kyseliny (polyunsaturated fatty acids)
RONS – reaktivní kyslíkaté a dusíkaté sloučeniny (reactive oxygen and nitrogen substances)
Rv – resolviny
SCFA – mastné kyseliny s krátkým řetězcem (short chain fatty acids)
SFA – nasycené mastné kyseliny (saturated fatty acids)
sn – stereospecific numbering
SREBP – vazebná bílkovina pro sterol regulující elementy (sterol regulatory element binding protein)
TG – triglycerid
TM – transmembránový
TNF-α – tumor nekrotizující faktor α
TX – tromboxany
VCAM – adhezivní molekuly vaskulárního endotelu (vascular cell adhesion molecule)
VLCFA – mastné kyseliny s velmi dlouhým řetězcem (very long chain fatty acids)
VLDL – lipoprotein o velmi nízké hustotě (very low density lipoprotein)
Práce byla podporována výzkumným záměrem MSM 0021620820 a MSM 0021620807.
Adresa pro korespondenci:
RNDr. Eva Tvrzická, CSc.
IV. interní klinika 1. LF UK a VFN
U Nemocnice 2, 128 08 Praha 2
fax: +420 224 923 524, e-mail: eva.tvrzicka@vfn.cz
Zdroje
1. Nelson DL Cox, MM. Lipid Biosynthesis. In: Principles of Biochemistry. New York: W.H. Freeman and Company 2005; 787–815.
2. Nelson DL, Cox MM. Lipids. In: Principles of Biochemistry. New York: W.H. Freeman and Company 2005; 343–363.
3. Havel RJ, Eder HA, Bragdon JH. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J Clin Invest 1955; 34: 1345–1355.
4. Smith WL. Nutritionally essential fatty acids and biologically indispensable cyclooxygenases. Trends Biochem. Sci 2008; 33: 27–37.
5. Berdyshev EV, Boichotb E, Lagenteb V. Anandamide – a new look on fatty acid ethanolamides. J Lipid Mediat Cell Signal 1996; 15: 49–67.
6. Praticė D, Lawson JA, Rokach J, et al. The isoprostanes in biology and medicine. Trends Endocrinol Metab 2001; 12: 243–247.
7. Haeggström JZ, Wetterholm A. Enzymes and receptors in the leukotriene cascade. Cell Mol Life Sci 2002; 59: 742–753.
8. Resh MD. Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochim Biophys Acta 1998; 1451: 1–16.
9. Burr ML, Fehily AMI, Gilbert JF, et al. Effects of changes in fat, fish, and fibre intake on death and myocardial reinfarction: diet and reinfarction trial (DART). Lancet 1989; II: 757–761.
10. Lorgeril de M, Salen P, Martin JL, et al. Mediterranean diet, traditional risk factors and the rate of cardiovascular complications after myocardial infarction. Final report of the Lyon Diet Heart Study. Circulation 1999; 99: 779–785.
11. GISSI-Prevenzione Investigators: Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: results of the GISSI-Prevenzione trial. Lancet 1999; 354: 447–455.
12. Chapkin RS, McMurray DN, Davidson LA, et al. Bioactive dietary long-chain fatty acids: emerging mechanisms of action. Br J Nutr 2008; 20: 1–6.
13. Montecucco F, Steffens S, Mach F. Insulin resistance: a proinflammatory state mediated by lipid-induced signaling dysfunction and involved in atherosclerotic plaque instability. Mediators Inflamm 2008; doi: 10.1155/2008/767623.
14. Lapillone A, Clarke SD, Herd WC. Polyunsaturated fatty acids and gene expression. Curr Opin Clin Nutr Metab Care 2004; 7: 151–156.
15. Pertwee RG. Ligands that target cannabinoid receptors in the brain: from THC to anandamide and betone. Addict Biol 2008; 13: 147–159.
16. Hannuksela ML, Liisanantti MK, Nissinen AE, et al. Biochemical markers of alcoholism. Clin Chem Lab Med 2007; 45: 953–961.
17. Chao W, Olson MS. Platelet-activating factor: receptors and signal transduction. Biochem J 1993; 292: 617–629.
18. Bennett MV, Verselis VK. Biophysics of gap junctions. Semin. Cell Biol 1992; 3: 29–47.
19. Lombardi F, Terranova P. Anti-arrhythmic properties of N-3 poly-unsaturated fatty acids (n-3 PUFA). Curr Med Chem 2007; 14: 2070–2080.
20. Brenner RR. Factors Influencing Fatty Acid Chain Elongation and Desaturation. In: Vergroesen AJ, Crawford M. eds. The role of fats in human nutrition. London: Academic Press 1989; 45–79.
21. Žák A, Jáchymová M, Tvrzická E, et al. The influence of polymorphism of – 493G/T MTP gene promoter and metabolic syndrome on lipids, fatty acids and oxidative stress. J Nutr Biochem 2008; 19: 634–641.
22. Žák A, Vecka M, Tvrzická E, et al. Composition of plasma fatty acids and non-cholesterol sterols in anorexia nervosa. Physiol Res 2005; 54: 443–451.
23. Burr GO, Burr MM. A new deficiency disease produced by the rigid exclusion of fat from the diet. J Biol Chem 1929; 82: 345–367.
24. Kinsella JE, Broughton S, Whelan JW. Dietary unsaturated fatty acids: interactions and possible needs in relation to eicosanoid synthesis. J Nutr Biochem 1990; 1: 123–141.
25. Crawford MA, Doyle W, William G, et al. The role of fats and EFAs for energy an cell structures in the growth of fetus and neonate. In: Vergroesen AJ, Crawford M. eds. The role of fats in human nutrition. London: Academic Press 1989; 82–116.
26. Seo T, Blanek WS, Deckelbaum RJ. Omega-3 fatty acids: molecular approaches to optima biological outcomes. Curr Opin Lipidol 2005; 16: 11–18.
27. Mead JF. In: Holman RT. ed. Progress in the Chemistry of Fats and Other Lipids. Oxford: Pergamon Press 1971; 161–189.
28. Dyerberg J. Linolenate-derived polyunsaturated fatty acids and prevention of atherosclerosis. Nutr Rev 1986; 44: 125–134.
29. Feskens EJ, Virtanen SM, Rasanen L, et al. Dietary factors determining diabetes and impaired glucose tolerance. A 20 year follow up of Finnish and Dutch Cohorts in Sevens Countries Study. Diabetes Care 1995; 18: 1104–1112.
30. Holman RT, Johnson SB, Hatch TE. A case of human linolenic acid deficiency involving neurological abnormalities. Am J Clin Nutr 1982; 35: 617–623.
31. Tvrzická E, Staňková B, Vecka M, Žák A. Mastné kyseliny. 1. Výskyt a biologický význam. Čas lék čes 2009; 148: 16–24.
32. Gadek JE, DeMichele SJ, Karlstad MD, et al. Effect of enteral feeding with eicosapentaenoic acid, γ-linolenic acid and antioxidants in patients with acute respiratory distress syndrome. Crit Care 1999; 27: 1409–1420.
33. Martinez M. The fundamentals and practice of docosahexaenoic acid therapy in peroxisomal disorders. Curr Opin Clin Nutr Metab Care 2000; 3: 101–108.
34. Oberkofler H, Liu YM, Esterbauer H, et al. Uncoupling protein-2 gene: reduced mRNA expression in intraperitoneal adipose tissue of obese humans. Diabetologia 1998; 41: 940–946.
35. Kenler AS, Swails WS, Driscoll DS, et al. Early enteral feeding in postsurgical cancer patients. Fish Oil Structured Lipid Based polymeric Formula Versus a Standard Polymeric Formula. Ann Surg 1996; 223: 316–333.
36. Clarke SD. The multi-dimensional regulation of gene expression by fatty acids: polyunsaturated fats as nutrient sensors. Curr Opin Lipidol 2004; 15: 13–18.
37. Ferdinandusse S, Denis S, Mooijer PA, et al. Identification of the peroxisomal beta-oxidation enzymes involved in the biosynthesis of docosahexaenoic acid. J Lipid Res 2001; 42: 1987–1995.
38. Dunstan J, Mori TA, Barden A, et al. Fish oil supplementation in pregnancy modifies neonatal alergen–specific immune response and clinical outcomes in infants at high risk of atopy: a randomised controlled trial. J Allergy Clin Immunol 2003; 112: 1178–1184.
39. Mihrshahi S, Peat JK, Marks GB, et al. Childhood Asthma Prevention Study. Eighteen-month outcomes of house dust mite avoidance and dietary fatty acid modification in the Childhood Asthma Prevention Study (CAPS). J Allergy Clin Immunol 2003; 111: 162–168.
40. Colomer R, Moreno-Nogueira JM, García-Luna PP, et al. N‑3 fatty acids, cancer and cachexia: a systematic review of the literature. Br J Nutr 2007; 97: 823–831.
41. Smith HJ, Greenberg NA, Tisdale MJ. Effect of eicosapentaenoic acid, protein and amino acids on protein synthesis and degradation in skeletal muscle of cachectic mice. Br J Cancer 2004; 91: 408–412.
Štítky
Adiktológia Alergológia a imunológia Angiológia Audiológia a foniatria Biochémia Dermatológia Detská gastroenterológia Detská chirurgia Detská kardiológia Detská neurológia Detská otorinolaryngológia Detská psychiatria Detská reumatológia Diabetológia Farmácia Chirurgia cievna Algeziológia Dentální hygienistkaČlánok vyšiel v časopise
Časopis lékařů českých

- Metamizol jako analgetikum první volby: kdy, pro koho, jak a proč?
- Fixní kombinace paracetamol/kodein nabízí synergické analgetické účinky
- Tramadol a paracetamol v tlumení poextrakční bolesti
- Realita liečby bolesti v paliatívnej starostlivosti v Nemecku
Najčítanejšie v tomto čísle
- Alveolární echinokokóza – nově se objevující onemocnění?
- Mastné kyseliny 2. Fyziologický a klinický význam
- Šedá eminence v medicíně – štítná žláza: 2. část
- Přehled chirurgické léčby vitiliga