
Synergistic Roles of Eukaryotic Translation Elongation Factors 1Bγ and 1A in Stimulation of Tombusvirus Minus-Strand Synthesis
Host factors are recruited into viral replicase complexes to aid replication of plus-strand RNA viruses. In this paper, we show that deletion of eukaryotic translation elongation factor 1Bgamma (eEF1Bγ) reduces Tomato bushy stunt virus (TBSV) replication in yeast host. Also, knock down of eEF1Bγ level in plant host decreases TBSV accumulation. eEF1Bγ binds to the viral RNA and is one of the resident host proteins in the tombusvirus replicase complex. Additional in vitro assays with whole cell extracts prepared from yeast strains lacking eEF1Bγ demonstrated its role in minus-strand synthesis by opening of the structured 3′ end of the viral RNA and reducing the possibility of re-utilization of (+)-strand templates for repeated (-)-strand synthesis within the replicase. We also show that eEF1Bγ plays a synergistic role with eukaryotic translation elongation factor 1A in tombusvirus replication, possibly via stimulation of the proper positioning of the viral RNA-dependent RNA polymerase over the promoter region in the viral RNA template.These roles for translation factors during TBSV replication are separate from their canonical roles in host and viral protein translation.
Published in the journal:
Synergistic Roles of Eukaryotic Translation Elongation Factors 1Bγ and 1A in Stimulation of Tombusvirus Minus-Strand Synthesis. PLoS Pathog 7(12): e32767. doi:10.1371/journal.ppat.1002438
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1002438
Summary
Host factors are recruited into viral replicase complexes to aid replication of plus-strand RNA viruses. In this paper, we show that deletion of eukaryotic translation elongation factor 1Bgamma (eEF1Bγ) reduces Tomato bushy stunt virus (TBSV) replication in yeast host. Also, knock down of eEF1Bγ level in plant host decreases TBSV accumulation. eEF1Bγ binds to the viral RNA and is one of the resident host proteins in the tombusvirus replicase complex. Additional in vitro assays with whole cell extracts prepared from yeast strains lacking eEF1Bγ demonstrated its role in minus-strand synthesis by opening of the structured 3′ end of the viral RNA and reducing the possibility of re-utilization of (+)-strand templates for repeated (-)-strand synthesis within the replicase. We also show that eEF1Bγ plays a synergistic role with eukaryotic translation elongation factor 1A in tombusvirus replication, possibly via stimulation of the proper positioning of the viral RNA-dependent RNA polymerase over the promoter region in the viral RNA template.These roles for translation factors during TBSV replication are separate from their canonical roles in host and viral protein translation.
Introduction
Plus-stranded (+)RNA viruses recruit numerous host proteins to facilitate their replication and spread [1], [2]. Among the identified host proteins are RNA-binding proteins (RBPs), such as ribosomal proteins, translation factors and RNA-modifying enzymes [1]–[5]. The subverted host proteins likely affect several steps in viral RNA replication, including the assembly of the replicase complex and initiation of RNA synthesis. However, the detailed functions of recruited host RBPs in (+)RNA virus replication are known only for a small number of host factors [2], [6]–[8].
Tomato bushy stunt virus (TBSV) is model plant RNA virus coding for two replication proteins, p33 and p92pol, which are sufficient to support TBSV replicon (rep)RNA replication in a yeast (Saccharomyces cerevisiae) model host [9], [10]. p33 and p92pol are components of the membrane-bound viral replicase complex, which also contains the tombusviral repRNA serving not only as a template for replication, but also as a platform for the assembly of the viral replicase complex [11]–[13]. Recent genome-wide screens and global proteomics approaches with TBSV and a yeast host revealed a large number of host factors interacting with viral components or affecting TBSV replication. The identified host proteins are involved in various cellular processes, such as translation, RNA metabolism, protein modifications and intracellular transport or membrane modifications [14]–[17].
Various proteomics analyses of the highly purified tombusvirus replicase has revealed at least five permanent resident host proteins in the complex, including the heat shock protein 70 chaperones (Hsp70) [18]–[21], glyceraldehyde-3-phosphate dehydrogenase [4], pyruvate decarboxylase [21], Cdc34p E2 ubiquitin conjugating enzyme [4], [21], [22], eukaryotic translation elongation factor 1A (eEF1A) [23], [24] and two temporary resident proteins, Pex19p shuttle protein [25] and the Vps23p adaptor ESCRT protein [24], [26], [27]. The functions of several of these proteins have been studied in some detail [4], [17], [18], [19], [20].
The emerging picture from systems biology approaches is that eukaroyotic translation elongation factors (eEFs), such as eEF1A, play several roles during TBSV replication. Accordingly, eEF1A has been shown to facilitate the assembly of the viral replicase complex and stimulate the initiation of minus-strand synthesis by the viral RNA-dependent RNA polymerase (RdRp) [23], [24]. Another translation elongation factor identified in our genome-wide screens with TBSV is eukaryotic elongation factor 1Bgamma (eEF1Bγ) [15]. eEF1Bγ is an abundant, but not essential cellular protein, which is part of the eukaryotic translation elongation factor 1B complex also containing the eEF1Bα subunit in yeast and the eEF1Bα and eEF1Bδ subunits in metazoans [28].The eEF1B complex is the guanine nucleotide exchange factor for eEF1A, which binds and delivers aminoacyl-tRNA in the GTP-bound form to the elongating ribosome. Additional roles have been ascribed to eEF1Bγ in vesicle-mediated intracellular protein transport, RNA-binding, vacuolar protein degradation, oxidative stress, intermediate filament interactions and calcium-dependent membrane-binding [29], [30], [31].
In this paper, we characterize the function of eEF1Bγ in TBSV replication. Our approaches based on yeast and in vitro replication assays reveal that eEF1Bγ is a component of the tombusvirus replicase and binds to the 3′-end of the viral RNA. Using a cell-free replication assay, we define that eEF1Bγ plays a role by enhancing minus-strand synthesis by the viral replicase. The obtained data support the model that eEF1Bγ opens up a ‘closed’ structure at the 3′-end of the TBSV (+)RNA, rendering the RNA compatible for initiation of (-)-strand synthesis. Moreover, we find that eEF1Bγ and eEF1A play nonoverlapping functions to enhance (-)-strand synthesis. Altogether, the two translation factors regulate TBSV replication synergistically by interacting with different portions of the viral (+)RNA and the replication proteins.
Results
Deletion of eEF1Bγ inhibits TBSV RNA accumulation in yeast model host
eEF1Bγ is coded by TEF3 and TEF4 nonessential genes in yeast [32], [33]. Single deletion of TEF3(CAM1) or TEF4 reduced TBSV repRNA accumulation to ∼25% (Figure 1A, lanes 3–8), while deletion of both genes resulted in even more inhibition, supporting TBSV repRNA accumulation only at 15% level (lanes 9–11). Expression of eEF1Bγ (Tef4p) in tef4Δ yeast increased TBSV replication to ∼80%, demonstrating that the defect in TBSV repRNA replication in tef4Δ yeast can be complemented.Altogether, these data established that eEF1Bγ plays an important stimulatory role in TBSV replication.

Depletion of eEF1Bγ inhibits (-)-strand synthesis by the TBSV replicase in a cell-free extract
To obtain direct evidence on the involvement of eEF1Bγ in TBSV replication, we prepared cell-free extracts (CFE) from a yeast strain lacking the TEF4 gene or from wt yeast. These yeast extracts contained comparable amount of total proteins (Figure 1C, right panel). The CFE extracts were programmed with the TBSV (+)repRNA and purified recombinant p33 and p92pol obtained from E. coli. Under these conditions, the CFE supports the in vitro assembly of the viral replicase, followed by a single cycle of complete TBSV replication, resulting in both (-)-stranded repRNA and excess amount of (+)-stranded progeny [20], [34]. Importantly in the case of a translation factor, this assay uncouples the translation of the viral proteins from viral replication, which are interdependent during (+)RNA virus infections.
CFE obtained from tef4Δ yeast supported only 29% of TBSV repRNA replication when compared with the extract obtained from wt yeast (Figure 1C, lane 2 versus 4). These data demonstrate that Tef4p plays an important role in the activity of the viral replicase complex.
To test if the decrease in TBSV repRNA replication in vitro was due to reduced (+) or (-)-strand synthesis, we measured the replication products under non-denaturing versus denaturing conditions (Figure 1C). We found that the amount of dsRNA [representing the newly-synthesized 32P-labeled (-)RNA product hybridized with the input (+)RNA; lane 1, Figure 1C, see also ref. [23]] and the newly-synthesized (+)RNA both decreased by ∼3-fold in CFE obtained from tef4Δ yeast in comparison with those products in the wt CFE (lane 3). Since the ratio of dsRNA and ssRNA did not change much in the CFEs (Figure 1C), the obtained data are consistent with the model that Tef4p (eEF1Bγ) affects the level of (-)RNA production, which then leads to proportionately lower level of (+)RNA progeny.
Adding purified recombinant eEF1Bγ to CFE from tef4Δ yeast supported TBSV repRNA replication to similar extent as the CFE from wt yeast (i.e., containing wt eEF1Bγ, Figure 1D, lanes 3–6 versus 1–2), indicating that the recombinant eEF1Bγ can complement the missing Tef4p in vitro, when the same amount of p33 and p92pol was provided. Using large amount of eEF1Bγ in the CFE-based assay did not further increase TBSV repRNA replication (Figure 1D, lanes 3–4), suggesting that eEF1Bγ should be present in optimal amount during TBSV replication.
eEF1Bγ stimulates initiation of (-)RNA synthesis by a viral RdRp in vitro
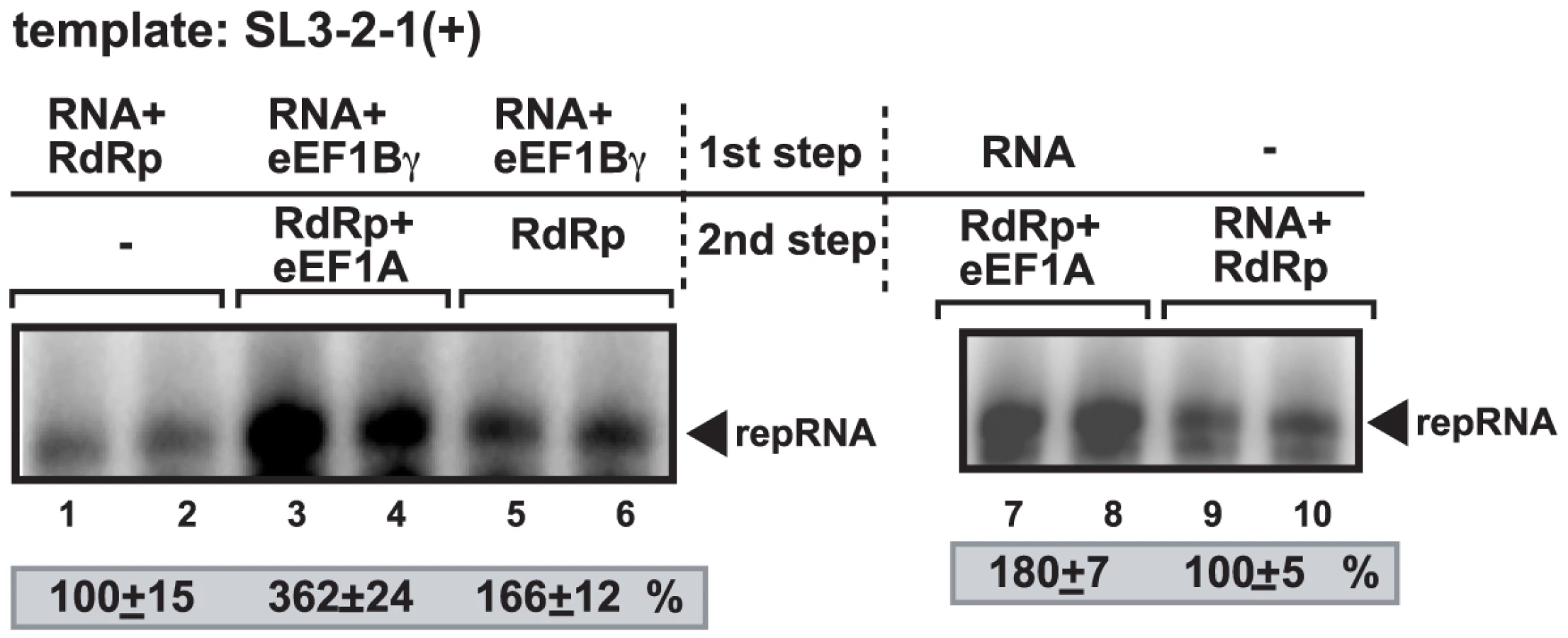
To obtain additional evidence if eEF1Bγ could stimulate RNA synthesis by the viral RdRp, we used the E. coli-expressed recombinant p88Cpol RdRp protein of Turnip crinkle virus (TCV). The TCV RdRp, unlike the E. coli-expressed TBSV p92pol or the closely-related Cucumber necrosis virus (CNV) p92pol RdRps, does not need the yeast CFE to be functional in vitro [35], [36]. Importantly, the template specificity of the recombinant TCV RdRp with TBSV RNAs is similar to the closely-related tombusvirus replicase purified from yeast or infected plants [10], [36], [37], [38]. The recombinant TCV RdRp preparation lacks co-purified eEF1Bγ (E. coli does not have a homolog), unlike the yeast or plant-derived tombusvirus replicase preparations, facilitating studies on the role of eEF1Bγ on the template activity of a viral RdRp. When we added various amounts of the highly purified recombinant eEF1Bγ to the TCV RdRp assay programmed with TBSV-derived SL3-2-1(+) RNA template, which is used by the TCV RdRp in vitro to produce the complementary (-)RNA product [37], we observed a ∼2-to-4-fold increase in (-)RNA synthesis by the TCV RdRp (Figure 2A, lanes 3–5). eEF1Bγ in the absence of the TCV RdRp did not give a 32P-labeled RNA product, excluding that our eEF1Bγ preparation contained RdRp activity (not shown). Altogether, our data suggest that eEF1Bγ can stimulate in vitro activity of TCV RdRp on a TBSV (+)RNA template, confirming a direct role for eEF1Bγ in viral (-)RNA synthesis by a viral RdRp.

To test if the stimulating activity of eEF1Bγ on the in vitro RdRp activity was due to binding of eEF1Bγ to the (+)RNA template and/or to the TCV RdRp protein, we performed assays, in which the recombinant eEF1Bγ was pre-incubated with the TCV RdRp or the (+)RNA template prior to the RdRp assay. These experiments revealed that pre-incubation of the purified eEF1Bγ with the TBSV-derived SL3-2-1(+) RNA template prior to the RdRp assay led to a ∼4.5-fold increase in (-)RNA products (Figure 2B, lanes 1–2). In contrast, pre-incubation of the TCV RdRp with the (+)RNA template (Figure 2B, lanes 3–4) or eEF1Bγ with the TCV RdRp (Figure 2B, lanes 7–8) prior to the RdRp assay did not result in increase in (-)RNA synthesis. Overall, data shown in Figure 2B imply that eEF1Bγ can stimulate (-)RNA synthesis only when eEF1Bγ binds to the (+)RNA template before the RdRp binding to the template.
To further test the stimulatory effect of eEF1Bγ, we also tested the RdRp activity in the presence of eEF1Bγ using a mutated (+)RNA template. The mutation [SL3-2-1m(+)] opens up the closed structure in the promoter region that leads to increased template activity [39]. The mutated template showed only ∼2-fold increased RNA products in the RdRp assay with eEF1Bγ (Figure 2C, lanes 3–4 versus 1–2). In contrast, eEF1Bγ did not stimulate RNA products when the negative-stranded RI-III(-) RNA was used as a template in the TCV RdRp assay (Figure 2C, lanes 9–10 versus 7–8). Thus, these data support the model that eEF1Bγ can mainly stimulate (-)-strand synthesis by the RdRp on the wt 3′ TBSV sequence, while it is not effective on the (-)RNA template.
eEF1Bγ binds to the 3′ end of the TBSV RNA in vitro
To test if eEF1Bγ directly binds to a particular region within the TBSV repRNA, we performed electrophoretic mobility shift (EMSA) experiments with purified eEF1Bγ and 32P-labeled regions of (+)repRNA that included known cis-acting elements involved in (-)RNA synthesis [39], [40], [41]. These experiments revealed that eEF1Bγ bound efficiently to the 3′-end of the TBSV (+)repRNA (construct SL3-2-1, carrying the terminal 3 stem-loop structures, Figure S1). Template competition experiments confirmed that SL3-2-1 RNA bound competitively to eEF1Bγ in vitro(Figure S1B).
To further define what sequence within SL3-2-1 is bound by eEF1Bγ, we used complementary DNA oligos to partially convert portions of SL3-2-1 into duplexes (RNA/DNA hybrids) as shown in Figure 3A. EMSA assay with purified recombinant eEF1Bγ revealed that the very 3′-terminal SL1 region had to be “free” (not part of the duplex) for eEF1Bγ to bind efficiently to the SL3-2-1 RNA (compare lane 1 with lane 5 in Figure 3A).

Since eEF1Bγ is known to bind to A-rich single-stranded sequences [32], we mutagenized the tetraloop (GAAA) sequence to either CUUG or GUUU tetraloop sequences (Figure 3B) that are expected to maintain the stability of the double-stranded stem. EMSA analysis showed that neither RNAs with the new tetraloop sequences bound efficiently to eEF1Bγ (Figure 3B, lanes 5–7 and 11–13). Based on the EMSA data, we conclude that the GAAA tetraloop region of SL1 is an efficient binding site for eEF1Bγ in vitro. However, we cannot exclude that eEF1Bγ binding may be dependent on stabilizing effects of the GNRA tetraloop on the stem structure. The loop nucleotides may or may not be involved in protein-RNA contacts.
Binding of eEF1Bγ to the 3′ end of the TBSV RNA is required for stimulation of (-)-strand RNA synthesis in vitro
To examine if binding of eEF1Bγ to SL1 is important for stimulation of (-)-strand RNA synthesis by the viral RdRp, we performed an in vitro RNA synthesis assay using a mutated SL3-2-1 carrying the ‘CUUG’ tetraloop instead of the wt ‘GAAA’ tetraloop sequence (Figure 4A). Unlike for the wt SL3-2-1 RNA, eEF1Bγ could not stimulate complementary RNA synthesis by the viral RdRp on the SL3-2-1cuug(+) template (Figure 4A, lanes 7–10 versus 1–4). These data suggest that binding of eEF1Bγ to the ‘GAAA’ tetraloop sequence of SL1 is important to stimulate (-)-strand synthesis by the viral RdRp in vitro.

eEF1Bγ co-purifies with the viral replicase complex and it binds to TBSV repRNA in yeast
To test if eEF1Bγ is a component of the tombusvirus replicase, we purified the His6-Flag-tagged p33 (HF-p33) replication protein via Flag-affinity purification from the detergent-solubilized membrane fraction of yeast [10]. We detected both p33 and eEF1Bγ in the purified preparation (Figure 5A, lane 1), suggesting that eEF1Bγ is likely part of the replicase complex [21]. Importantly, eEF1Bγ was not found in the control samples containing the His6-tagged p33 (H-p33) that were also purified via the Flag-affinity procedure (Figure 5A, lane 2). Since eEF1Bγ does not seem to bind to p33 or p92 replication proteins (data not shown), it is likely that eEF1Bγ was co-purified with p33 via the viral RNA template in the viral replicase complex.

To demonstrate that eEF1Bγ can indeed bind to the TBSV (+)repRNA in cells, we Flag-affinity-purified His6-Flag-tagged eEF1Bγ from the detergent-solubilized membrane fraction and also from the soluble (cytosolic) fraction of yeast. Interestingly, the viral RNA was co-purified with eEF1Bγ from both fractions (Figure 5B, lanes 3 and 7). These data confirmed that eEF1Bγ binds to the viral RNA in yeast.
Since eEF1Bγ was found in association with the TBSV repRNA in the cytosolic fraction of yeast, it is possible that eEF1Bγ might affect the viral RNA recruitment from the cytosol into replication that takes place on the peroxisomal or ER membrane surfaces [42], [43]. Therefore, we tested the recruitment of the TBSV (+)repRNA to the membrane fraction in our CFE assay [23]. We found that eEF1Bγ did not facilitate the association of the TBSV (+)repRNA with the membrane when applied in the absence of p33/p92 replication proteins (Figure S2). Moreover, eEF1Bγ did not further increase the amount of TBSV (+)repRNA bound to the membrane in the presence of p33/p92 replication proteins, which are needed for RNA recruitment (Figure S2, lanes 3–4 and 8–10) [24]. Therefore, we conclude that eEF1Bγ is unlikely to promote the recruitment of the TBSV (+)repRNA to the membrane.
Synergistic effect of eEF1Bγ and eEF1A on the activity of the viral RdRp in vitro
Since both eEF1Bγ and eEF1A bind to the 3′-terminal region of the TBSV (+)RNA (Figure 3) and ref: [23], [24], it is possible that they could affect each other's functions during replication. To test the mutual effect of eEF1Bγ and eEF1A on the (-)-strand RNA production of the viral RdRp, we performed in vitro RdRp assays with purified eEF1A and recombinant eEF1Bγ as shown in Figure 6. Based on previous experiments, eEF1Bγ was known to stimulate (-)-strand synthesis the most when pre-incubated with the template (+)RNA (Figure 2B). In contrast, pre-incubation of eEF1A with the viral RdRp was more effective than pre-incubation of eEF1A with the template RNA [23]. Therefore, we performed the pre-incubation experiments prior to the RdRp assay as shown in Figure 6. We found the largest stimulation of (-)-strand synthesis by the viral RdRp in a dual pre-incubation assay, when eEF1Bγ was pre-incubated with the viral RNA template, while eEF1A was separately pre-incubated with the viral RdRp (Figure 6, lanes 3–4). Pre-incubation of eEF1Bγ with the viral RNA template (lanes 5–6) or pre-incubation of eEF1A with the viral RdRp (lanes 7–8) were about half as efficient in stimulation of (-)-strand synthesis than the dual pre-incubation assay (lanes 3–4). Therefore, these data support the model that eEF1Bγ and eEF1A both promote (-)-strand synthesis and their effect is synergistic, likely involving separate mechanisms (see Discussion).
Silencing of eEF1Bγ in plants inhibits TBSV RNA accumulation
To obtain evidence on the importance of eEF1Bγ in TBSV replication in the natural plant hosts, we knocked down the expression of the eEF1Bγ gene in Nicotiana bethamiana leaves via VIGS (virus-induced gene silencing). Efficient knocking down of eEF1Bγ mRNA level in N. benthamiana (Figure 7B) only resulted in slightly reduced growth of the plants without other phenotypic effects (Figure 7A). The accumulation of TBSV genomic RNA, however, was dramatically reduced in both inoculated (Figure 7B, lanes 1–5) and the systemically-infected young leaves (Figure 7C, lanes 1–4) when compared with the control plants infected with the ‘empty’ Tobacco rattle virus (TRV) vector. The lethal necrotic symptoms caused by TBSV in N. benthamiana were also greatly attenuated in the eEF1Bγ knock-down plants (Figure 7A). Therefore, we conclude that eEF1Bγ is essential for TBSV genomic RNA accumulation in N. bethamiana.

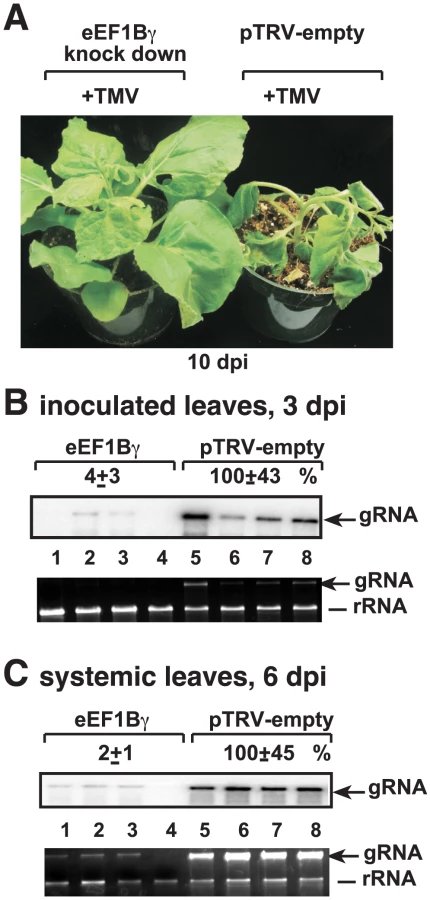
Silencing of eEF1Bγ in plants inhibits Tobacco mosaic virus RNA accumulation
To test if eEF1Bγ is also needed for the replication of other plant RNA viruses, we infected eEF1Bγ-silenced N. benthamiana leaves with the unrelated Tobacco mosaic virus (TMV) RNA (Figure 8A). We found that the severe symptoms caused by TMV were greatly ameliorated in eEF1Bγ knock-down plants (Figure 8A). Accumulation of TMV genomic RNA was also dramatically reduced in both inoculated (Figure 8B) and systemically-infected (Figure 8C) leaves of the eEF1Bγ knock-down plants. Based on these data, eEF1Bγ seems to be needed for TMV replication and/or spread in plants. Thus, our data have revealed new functions for eEF1Bγ in plant RNA virus replication and spread.
Discussion
Tombusviruses, similar to other (+)RNA viruses, subvert a yet unknown number of host-coded proteins to facilitate robust virus replication in infected cells. The co-opted host proteins could be part of the viral replicase complexes and provide many yet undefined functions. Translation factors, such as eEF1Bγ and eEF1A, are among the most common host factors recruited for (+)RNA virus replication [23], [24]. While eEF1A is an integral component of the tombusvirus replicase complex [23], [24] and several other viral replicases [44], [45], [46], the function of eEF1Bγ in tombusvirus replication is studied in this paper. Co-purification experiments with the p33 replication protein, which is the most abundant protein component in the tombusvirus replicase complex [21], [22], revealed that eEF1Bγ is a permanent member of the replicase (Figure 5A). eEF1Bγ is likely recruited into the viral replicase via the viral (+)RNA, which is bound to eEF1Bγ in both cytosolic and membranous fractions (Figure 5B). The possible role of host proteins or membrane lipids in assisting the recruitment of eEF1Bγ for TBSV replication cannot be excluded. Accordingly, eEF1Bγ has been shown to bind to a large number of host proteins (www.yeastgenome.org). For example, eEF1A, which is also a permanent member of the tombusvirus replicase, is known to interact with eEF1Bγ [47], [48], [49] and eEF1A might facilitate the recruitment of eEF1Bγ and possibly other translation factors. The binding of eEF1Bγ to intracellular membranes has also been shown before [32]. Altogether, our model predicts that the viral (+)RNA could be involved in recruitment of eEF1Bγ into viral replication (Figure 5). However, the opposite model that eEF1Bγ facilitates the recruitment of the TBSV (+)RNA into replication is not supported by our in vitro data (Figure S2). Indeed, addition of eEF1Bγ to the CFE assay did not increase the membrane-bound fraction of TBSV (+)repRNA in the absence or presence of the viral replication proteins (Figure S2).
eEF1Bγ selectively enhances minus-strand synthesis by opening the closed 3′-terminus during TBSV RNA replication
We confirmed a direct role for eEF1Bγ in RNA synthesis in vitro by using a cell-free extract prepared from tef4Δ yeast that supported (-)-strand RNA synthesis ∼3-fold less efficiently than CFE from wt yeast (Figure 1). Moreover, in vitro assays with highly purified eEF1Bγ and the recombinant TCV RdRp, which is closely homologous with the TBSV p92pol, also revealed that eEF1Bγ stimulates (-)-strand synthesis by binding to the viral (+)RNA template (Figure 3). Accordingly, pre-incubation of eEF1Bγ and the TBSV-derived template RNA prior to the RdRp assay led to the highest level of stimulation of (-)RNA synthesis (Figure 2). On the other hand, eEF1Bγ does not stimulate the RdRp activity directly, since pre-incubation of eEF1Bγ with the RdRp did not lead to more efficient (-)-strand RNA synthesis in vitro (Figure 2). We propose that eEF1Bγ modifies the structure of the (+)-strand template prior to initiation of (-)-strand synthesis that leads to more efficient RNA synthesis as described below.
In vitro initiation of (-)-strand synthesis by the viral RdRp requires the gPR promoter consisting of a short 3′-terminal single-stranded tail and a stem-loop (SL1) sequence [39], [50]. However, the gPR region is present in a ‘closed’ structure in the TBSV (+)RNA due to base-pairing of a portion of the gPR with the RSE present in SL3 as shown in Figure 9. This interaction makes the TBSV (+)RNA poor template in the in vitro assay due to the difficulty for the viral RdRp to recognize and/or open the ‘closed’ structure [39]. Our current work with eEF1Bγ, however, suggests that eEF1Bγ can bind to the tetraloop region of SL1 (and to an A-rich sequence in SL2) that leads to melting of the base-paired structure and opening the stem of SL1 and the RSE-gPR base-pairing as shown schematically in Figure 9B. We propose that the open structure can be recognized efficiently by the viral replicase leading to efficient initiation of (-)-strand synthesis (Figure 9B). This model is supported by several pieces of evidence presented in this paper, including (i) stimulation of (-)-strand synthesis by eEF1Bγ when the wt SL1 is present in the template; (ii) lack of stimulation of(-)-strand synthesis by eEF1Bγ when a mutated SL1 (tetraloop mutant), which does not bind efficiently to eEF1Bγ, was used as a template in the in vitro assay; (iii) stimulation of (-)-strand synthesis when eEF1Bγ was pre-incubated with the (+)-strand template, but not when eEF1Bγ was pre-incubated with the viral RdRp (Figure 2); and (iv) the lack of stimulation of (+)-strand synthesis on a (-)-strand template by eEF1Bγ (Figure 2). In addition, eEF1Bγ stimulated (-)-strand synthesis by the viral RdRp when a partially complementary RNA oligo was hybridized with the SL1 region (Figure 4B). However, eEF1Bγ could not efficiently bind to the 3′-end of the TBSV RNA when it formed a hybrid (duplex) with a perfectly complementary DNA oligo (Figure 3A), suggesting that eEF1Bγ can melt only the local secondary structure, but cannot unwind more extended duplex regions. An alternative possibility is that eEF1Bγ protein stabilizes the unpaired structure (when the SL1 structure is kinetically pairing/unpairing), rather than implying that it actively "opens" the structure.

An intriguing aspect of our model is the possible regulation of the “open” and “closed” structure of the 3′ UTR by eEF1Bγ. Displacement of eEF1Bγ bound to the 3′-end by the viral replicase during (-)-strand synthesis could make the 3′-terminus of the (+)-strand RNA fold back into a ‘closed’ structure. This could prevent efficient re-utilization of the original (+)-strand template during TBSV replication, and the switch to efficient (+)-strand synthesis on the (-)RNA intermediate (Figure 9B). This model can also explain why the newly made (+)-strand RNA progeny will not enter the replication cycle in the absence of bound eEF1Bγ within the originally-formed replicase complexes as observed previously in the CFE assay [20]. We propose that the new (+)RNA progeny need to leave the replicase complex, then bind to eEF1Bγ in the cytosol and assemble new replicase complexes, followed by a new round of viral RNA replication. Thus, this model suggests that eEF1Bγ plays a key role in regulation of the use of (+)-strand RNAs in TBSV replication (Figure 9B).
Our finding of TBSV RNA binding by eEF1Bγ adds to the growing list of RNAs bound by eEF1Bγ. For example, the 3′ UTR of vimentin mRNA is bound by eEF1Bγ [51], which led the authors to suggest that eEF1Bγ plays a role in vimentin mRNA subcellular localization by also binding to cytoskeleton or membranes. eEF1Bγ also binds to the tRNA-like structure at the 3′ UTR of BMV, albeit the relevance of this binding is currently unclear [51]. Also, the actual role of eEF1Bγ in the VSV replicase is currently not defined [31].
Translation elongation factors seem to be important for replication of many RNA viruses. For example, EF-Tu and EF-Ts play a role in replication of bacteriophage Qbeta [52], [53]. The eukaryotic homolog of EF-Tu, eEF1A was found to bind to viral RNAs, such as TBSV, Turnip yellow mosaic virus (TYMV) [54], West Nile virus (WNV), Dengue virus, hepatitis delta virus, TMV, Brome mosaic virus, and Turnip mosaic virus [55], [56], [57], [58], [59], [60] and to viroid RNAs [61]. Therefore, it is highly probable that many (+)-strand RNA viruses recruit translation elongation factors to facilitate and regulate their replication in infected cells.
Nonoverlapping roles of eEF1Bγ and eEF1A in stimulation of (-)-strand synthesis
The emerging picture on the functions of eEF1Bγ and eEF1A is that these translation elongation factors play different, yet complementary roles in TBSV replication as suggested in Figure 9B. While eEF1Bγ binds to SL1, eEF1A has been shown to bind to both p92pol RdRp and the SL3 region of TBSV (+)repRNA [23], [24]. The binding of the RNA by eEF1Bγ promotes the opening of the closed 3′-terminal structure, whereas eEF1A facilitates the proper and efficient binding of the RdRp to the 3′ terminal RSE sequence of the viral RNA, which is required for the assembly of the viral replicase complex [11], [39], prior to initiation of (-)-strand synthesis (Figure 9) [23], [24]. The binding of eEF1A-RdRp complex to the RSE might lead to proper positioning of the RdRp over the 3′-terminal gPR promoter sequence opened up by eEF1Bγ, thus facilitating the initiation of (-)RNA synthesis starting from the 3′-terminal cytosine (Figure 9B). Altogether, the two translation factors facilitate the efficient initiation of (-)-strand synthesis in addition to reducing the possibility of re-utilization of the (+)-strand template for additional rounds of (-)-strand synthesis. This regulation of RNA synthesis by the co-opted host factors shows the specialized use of host components to serve the need of viral replication.
eEF1Bγ is required for TBSV replication in yeast and plant hosts
The current work also provides evidence that eEF1Bγ is a key factor in TBSV replication in yeast (Figure 1) and in N. benthamiana (Figure 7). Since eEF1Bγ is a highly conserved protein in all eukaryotes [32], it is not surprising that yeast eEF1Bγ, similar to the plant eEF1Bγ, can be co-opted for TBSV replication. Interestingly, deletion of either TEF3 or TEF4 genes reduced TBSV repRNA accumulation in yeast, suggesting that eEF1Bγ is present in limiting amount or eEF1Bγ is present in not easily accessible forms (in protein complexes) and/or locations in yeast cells. Silencing of eEF1Bγ in N. bethamiana showed even more inhibition of TBSV RNA accumulation than deletion of eEF1Bγ genes in yeast. This is likely due to the robust antiviral response (i.e., induced gene silencing) of the plant host, which could result in degradation of the small amount of viral RNA produced by the less efficient viral RNA replication in the presence of limited eEF1Bγ in the knock-down plants.
Silencing of eEF1Bγ expression in N. benthamiana also reduced the accumulation of the unrelated TMV (Figure 8), which belongs to the alphavirus-like supergroup. These data suggest that eEF1Bγ is likely involved in TMV replication, which also contains a highly structured 3′- end [54]. Therefore, it is possible that eEF1Bγ is co-opted by different plant RNA viruses, and possibly other RNA viruses as well.
Conclusion
Overall, the current work suggests three major functions for eEF1Bγ in TBSV replication (Figure 9): (i) enhancement of the minus-strand synthesis by opening the ‘closed’ 3′-end of the template RNA; (ii) reducing the possibility of re-utilization of (+)-strand templates for repeated (-)-strand synthesis; and (iii) in coordination with eEF1A, stimulation of the proper positioning of the viral RdRp over the promoter region in the viral RNA template. These roles for eEF1Bγ and eEF1A are separate from their canonical roles in host and viral protein translation.
Materials and Methods
Yeast strains and expression plasmids
Saccharomyces cerevisiae strain BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0) and the single-gene deletion strain of the TEF4-encoded form of eEF1Bγ (tef4Δ) were obtained from Open Biosystems (Huntville, AL). TKY680 strain in which both yeast encoded eEF1Bγ, TEF4 and TEF3 were deleted (MATa ura3-52 leu2Δ1 his3Δ200 trp1Δ101 lys2-801 tef3::LEU2 tef4::TRP1) and its isogenic wild type TKY677 (MATa ura3-52 leu2Δ1 his3Δ200 trp1Δ101 lys2-801) as well as the isogenic single deletion mutant strains, TKY678 (MATa ura3-52 leu2Δ1 his3Δ200 trp1Δ101 lys2-801 tef3::LEU2) and TKY 679 (MATa ura3-52 leu2Δ1 his3Δ200 trp1Δ101 lys2-801 tef4::TRP1) were published previously [30]. The following plasmids pESC-GAL1-Hisp33/GAL10-DI-72, pGAD-CUP1-p92 pYES-GAL1-p92, pCM189-TET-His92 were described earlier [21], [22]. URA3 based pGBK-ADH- Hisp33/GAL1-DI72, pGBK-CUP1-HisFLAGp33/GAL1-DI-72, and pGBK-CUP1- Hisp33/GAL1-DI-72 plasmids were constructed by Daniel Barajas (unpublished result). The URA3 based, low copy-number plasmid, pYC-GAL1-Tef4 expressing non-tagged full-length Tef4 protein was constructed as follows: pYC/NT-C plasmid was digested with BamHI and XhoI restriction enzymes and then PCR product of the TEF4 gene was generated with primers #2089 (ccgcGGATCCATGTCCCAAGGTACTTTATAC) and #2320 (CGCCTCGAGTTATTTCAAAACCTTACCGTCAACAATTTCC) and digested with the same restriction enzymes, followed by ligation. The plasmid pYES-NTC2-GAL1-HisTef4 expressing His6-tagged Tef4p protein was created with the same restriction enzymes using pYES-NT-C2.
HIS3-based pEsc-His/Cup-FLAG plasmid [20] was digested with BamHI and XhoI restriction enzymes and then PCR product of the TEF4 gene was generated with primers #2089 and #2320 and digested with the same restriction enzymes, followed by ligationto obtain pEsc-His/Cup-FLAG-TEF4.
In vivo replication assay
HIS3 based pESC-GAL1-His33/GAL10-DI-72 and LEU2 based pGAD-CUP1-Hisp92 plasmids were transformed into tef4Δ strain. In the in vivo complementation assay, non-tagged Tef4p protein was expressed from URA3 plasmid pYC-GAL1-Tef4 and TEF4 mRNA was detected with a specific probe generated by the T7 transcription of the PCR product obtained with primers #2089 and #3788 (TAATACGACTCACTATAGGATTATTTCAAAACCTTACCGTCAACAATTTCC).
TKY680 (tef3Δ/tef4Δ), the isogenic TKY679 (tef4Δ), TKY678 (tef3Δ) and wild type TKY677 yeast were transformed with plasmids pESC-GAL1-His33/GAL10-DI-72 and pCM189-TET-His92. Yeast was pre-grown at 23°C overnight in 3 ml synthetic complete dropout medium lacking the relevant amino acids containing 2% glucose and 1 mg/ml doxycyclin to suppress p92 expression by the inhibition of TET promoter and then TBSV replication was launched by replacing the media with 2% galactose without doxycycline. Cells were harvested at 48 h time point. Total RNA extraction from yeast cells and Northern blotting and Western blotting were done as previously described [15], [24].
Expression and purification of recombinant eEF1Bγ protein
pEsc-His/Cup-FLAG-TEF4 plasmid was transformed into tef4Δ strain. Yeast was pre-grown overnight at 29°C in 2 ml synthetic complete dropout medium lacking histidine (SC-H- medium) containing 2% glucose. The volume of the media was increased up to 100 ml 16 h later and copper sulfate was added to a final concentration of 50 µM for induction of protein expression. Yeast was grown to 0.8 OD600 (∼4–6 h). Then, yeast cells were harvested and broken by glass beads in a FastPrep cell disruptor followed by Flag-affinity purification of FLAG-Tef4p protein [34]. The bacterial heterologous expression and purification of His6-tagged Tef3 protein from plasmid pTKB523 was performed as described in ref: [62] using only the Ni affinity column step.
Tombusvirus replication assay using yeast cell free extract
Yeast extract capable of supporting TBSV replication in vitro was prepared as described [20]. The newly synthesized 32P-labeled RNA products were separated by electrophoresis in a 5% polyacrylamide gel (PAGE) containing 0.5x Tris-borate-EDTA (TBE) buffer with 8 M urea. To detect the double-stranded RNA (dsRNA) in the cell-free replication assay, the 32P-labeled RNA samples were divided into two aliquotes: one half was loaded onto the gel without heat treatment in the presence of 25% formamide, while the other half was heat denatured at 85°C for 5 min in the presence of 50% formamide [20].
To test the in vitro activity of Tef4p, different concentrations (26 and 13 pmol) of purified FLAG/His6-Tef4p was added to 0.25 µg (4 pmol) DI-72 (+)repRNA transcript and incubated in the presence of yeast cell-free extract and reaction buffer for 10 minutes at RT followed by the addition of MBP-p33 and MBP-p92 along with the rest of the reaction components. The reaction was performed at 25°C for 3 h and analyzed as above.
In vitro TCV p88C RdRp assay
The TCV RdRp reactions were carried out as previously described for 2 h at 25°C [36], except using 7 pmol template RNA and 2 pmol affinity-purified MBP-p88C. Different concentrations of eEF1Bγ (6xHis-affinity purified recombinant Tef3p obtained from E. coli or Flag-affinity purified HF-Tef4p obtained from yeast) were added to the reaction at the beginning or as indicated in the text and Figure 2. legend. The 32P-labeled RNA products were analyzed by electrophoresis in a 5% PAGE/8 M urea gel [63]. The 86-nt 3′ noncoding region of TBSV genomic RNA and its mutants were used as the template in the RdRp assay [24], [36]. RNA templates were generated with T7 transcription using PCR products obtained with the following primers: #1662 (TAATACGACTCACTATAGGACACGGTTGATCTCACCCTTC) and #1190 (GGGCTGCATTTCTGCAATG) for SL3-2-1(+), #1662 and #4390 (GGGCTGCACAAGTGCAATGTTCCGGTTGTCCGGT) for SL3-2-1cuug(+). SL3-2-1m(+) RNA was generated with T7 transcription on PCR products amplified with primers #1662 and #1190, on a plasmid template harboring GGGCU nucleotide-deletion in SL3 region as described [39]. A duplex RNA was generated by hybridizing SL3-2-1(+) and SL3-2-ds1(-) made by T7 transcription of the PCR product using primers #4361 (GTAATACGACTCACTATAGGGCTACTTCCGGTTGTCCGGTAGTGCTTCC) and # 4362 (CGGTTGATCTGACCCTTCGG). For hybridization, equal amounts of both RNAs were mixed in 1X STE buffer [0.1 M NaCl 10 mM Tris-HCl (pH 8.0) 1 mM EDTA (pH 8.0)] followed by treatments: 94°C for 15 s, 70 cycles with gradually lowering the temperature by 1°C at each cycle for 30 s and finally 20°C for 30s.
Gel mobility shift assay (EMSA) with eEF1Bγ
For EMSA, 6xHis-Flag tagged Tef4p was purified from a yeast tef4Δ strain with anti-FLAG M2-agarose affinity resin. Different concentrations (0.6, 0.5 and 0.4 pmol) of HF-Tef4p protein was used for incubation with 0.2 pmol of 32P-labeled SL3/2/1(+) RNA or mutated RNAs at 25°C in a binding buffer [50 mM Tris-HCl (pH 8.2), 10 mM MgCl2, 10 mM DTT, 10% glycerol, 2 U of RNase inhibitor (Ambion)]. Samples were incubated at 25°C for 15 min, then resolved in 4% nondenaturing polyacrylamide gel [23]. Similar experiments were also performed with 6xHis-affinity purified recombinant Tef3p obtained from E. coli (not shown).
Flag-affinity purification of eEF1Bγ-TBSV repRNA complex
For the co-purification of TBSV DI-72 repRNA and eEF1Bγ protein, the yeast tef4Δ strain was co-transformed with pGBK-ADH-Hisp33/GAL1-DI72, pGAD-CUP1-Hisp92 and pESC-CUP1-HisFLAG-Tef4. The pESC-CUP1-FLAGHis-Tef4 plasmid was replaced with the pESC plasmid in the control experiment. Yeast was pre-grown overnight at 29°C in 2 ml SCULH- medium containing 2% glucose and 5 µM copper sulfate. The volume of the media was increased to 20 ml after 16 h for an additional 10 h (OD600 of ∼0.8), then the cultures were transferred to 20 ml SCULH- medium containing 2% galactose to induce TBSV DI-72 RNA transcription at 23°C. The transcription of DI-72 RNA was stopped by changing to the media containing 2% glucose after 8 h. The cultures were diluted to 200 ml and copper sulfate was added to a final concentration of 50 µM to induce the expression of Flag-tagged Tef4 protein. After incubation at 23°C for 24 h, the samples were centrifuged at 3000 rpm for 4 min. Cells (∼1 g) were re-suspended in 2 ml TG Buffer (50 mM Tris–HCl [pH 7.5], 10% glycerol, 15 mM MgCl2, and 10 mM KCl) supplemented with 0.5 M NaCl and 1% [V/V] YPIC yeast protease inhibitor cocktail (Sigma) and RNase inhibitor (Ambion). Yeast cells were broken by glass beads in a FastPrep cell disruptor (MP Biomedicals) 4 times for 20 sec each at speed 5.5. Samples were removed and incubated 1 min in an ice-water bath after each treatment. The samples were centrifuged at 500 ×g for 5 min at 4°C to remove glass beads, unbroken cells and debris then supernatant was moved into fresh pre-chilled tubes. After being centrifuged again at 500 ×g for 5 min at 4°C supernatant transferred into fresh pre-chilled tubes and soluble (SU) and membrane (ME) fractions containing the viral replicase complex were separated with centrifugation at 35,000 ×g for 15 min at 4°C. The SU fraction was applied on 0.1 ml anti-FLAG M2-agarose affinity resin (Sigma) and Tef4 protein tagged with 6xHis- and FLAG affinity tags was purified. Before applying ME fraction on the anti-FLAG M2 resin, solubilization of the membrane-bound replicase was performed in 1 ml TG buffer with 0.5 M NaCl, 1% [V/V] YPIC yeast protease inhibitor cocktail (Sigma), and 2% Triton X-100 via rotation for 2 hours at 4 °C. The solubilized membrane fraction was centrifuged at 35,000 ×g at 4°C for 15 min and the supernatant was added to the resin pre-equilibrated with TG buffer supplemented with 0.5 M NaCl and 0.5% Triton X-100, followed by gentle rotation for 2 h at 4°C. The unbound proteins were removed by gravity flow, and the resin was washed two times with 1 ml TG buffer supplemented with 0.5 M NaCl, 0.5% Triton X-100 and once with 1 ml TG buffer, 0.5% Triton without NaCl. The bound proteins were eluted with 150 µl TG buffer without NaCl, 0.5% Triton X-100, supplemented with 150 µg/ml flag peptide and 1% yeast protease inhibitor cocktail via gentle tapping the column occasionally for 2 h at 4°C. After centrifugation at 600 ×g 2 min at 4°C, semi-quantitative RT-PCR was performed to detect TBSV repRNA co-purified with eEF1Bγ using primers, #359 (GTAATACGACTCACTATAGGAAATTCTCCAGGATTTC) and #1190, amplifying full length (+)repRNA.
Purification of the viral replicase
To test if eEF1Bγ is present in the viral replicase, yeast tef4Δ strain was transformed with pGBK-CUP1-HisFLAGp33/GAL1-DI-72, pGAD-CUP1-Hisp92 and pYES-GAL1-HisTef4. In the control experiment, 6xHisp33was expressed from pGBK-CUP1-Hisp33/GAL1-DI-72. Yeast cultures were grown in SC-ULH- media containing 1% raffinose and 1% galactose with 5 µM copper-sulfate for 4 days with increasing the volume of the culture from 2 ml to 100 ml to a final OD600 of∼ 1.0. After harvesting of cells, co-purification of 6xHis-tagged Tef4p with HF-p33 (part of the viral replicase) was conducted by using anti-FLAG M2-agarose affinity resin as described above (in the section: FLAG-affinity purification of eEF1Bγ-TBSV repRNA complex), with the exception that only solubilized ME fraction was loaded on the column. Proteins bound to affinity resin were eluted by incubation with 150 µl buffer containing FLAG peptide and precipitated with Trichloroacetic acid (TCA) [64]. Samples were analyzed by SDS-PAGE and Western blotting.
Virus induced gene silencing of eEF1Bγ in N. benthamiana plants
Virus-induced gene silencing (VIGS) in N. benthamiana was done as described [65], [66]. To generate the VIGS vector (pTRV2- eEF1BγNt), a 314-bp cDNA fragment of NteEF1Bγ was RT-PCR amplified from a total RNA extract of N. benthamiana using the following pair of primers: #2993 (CGCGGATCCAAAGGTTTCTGGGACATGTATGA) and #2994 (CGCCTCGAGACACGCTCCTTCTGTGATTCATC) and inserted into the corresponding (BamHI/XhoI) restriction sites of pTRV2 plasmid.
The sequence of the N. tabacum eEF1Bγ gene (GenBank: ACB72462.1) was derived via a BLASTP search based on the C- terminal (translation elongation factor) domain (aa 252–412) of the Saccharomyces cerevisie Tef4 protein. The selected sequence (TC64920) from the Solanaceae Genomics Resource (www.tigr.org) gave 98% identity with N. tabacum EF1Bγ -like gene (GB#: EU580435.1).
To confirm the silencing of the EF1Bγ gene in N. benthamiana, we performed RT-PCR amplification with primer pairs: #2952 (CGCGGATCCGGAAAGGTTCCTGTGCTTGA) and #2992 (CGCCTCGAGGTCCAGAAGTATCTCTCTACATGTGG) on total RNA extract of pTRV2- EF1BγNt and pTRV2empty agro-infiltrated N benthamiana plants. PCR conditions were as follows: 27 cycles of 94°C 20sec, 60°C 30sec, 68°C 30 sec with HiFi Taq polymerase. Tubulin mRNA control from the same total RNA samples was detected by RT-PCR using primers #2859 (TAATACGACTCACTATAGgaACCAAATCATTCATGTTGCTCTC) and #2860 (TAGTGTATGTGATATCCCACCAA) [65]. The leaves of VIGS-treated plants were sap inoculated with TBSV, or TMV on the 9th day after silencing [65]. Total RNA was extracted 3 or 5 days post inoculation [65]. For Northern blot analysis of the viral RNA level, we prepared 32P-labeled complementary RNA probes specific for the 3′-ends of the viral genomic RNAs based on T7 transcription. To obtain the PCR templates for the probes, we used the following primers for TBSV: #1165 (AGCGAGTAAGACAGACTCTTCA) and #22; for TMV: #2890 (TCTGGTTTGGTTTGGACCTC) and #2889 (GTAATACGACTCACTATAGGGATTCGAACCCCTCGCTTTAT).
In vitro viral RNA recruitment assay
The TBSV viral RNA is recruited to the membrane from the soluble fraction with the help of TBSV replication proteins and host factors present in the yeast CFE. The in vitro RNA recruitment reaction was performed according to [20], [23], except that 32P-labeled DI-72 (+)repRNA were used and rCTP, rUTP, 32P-labeled UTP, and Actinomycin D were omitted from the assay. As a negative control, p33 and p92 were omitted from the reaction to detect DI-72 binding nonselectively to host proteins present in the membrane.
Supporting Information
Zdroje
1. NoueiryAOAhlquistP 2003 Brome Mosaic Virus RNA Replication: Revealing the Role of the Host in RNA Virus Replication. Annu Rev Phytopathol 41 77 98
2. NagyPD 2008 Yeast as a model host to explore plant virus-host interactions. Annu Rev Phytopathol 46 217 242
3. StraussJHStraussEG 1999 Viral RNA replication. With a little help from the host. Science 283 802 804
4. WangRYNagyPD 2008 Tomato bushy stunt virus Co-Opts the RNA-Binding Function of a Host Metabolic Enzyme for Viral Genomic RNA Synthesis. Cell Host Microbe 3 178 187
5. LiZNagyPD 2011 Diverse roles of host RNA binding proteins in RNA virus replication. RNA Biol 8 305 315
6. NagyPDPoganyJ 2006 Yeast as a model host to dissect functions of viral and host factors in tombusvirus replication. Virology 344 211 220
7. BrintonMA 2001 Host factors involved in West Nile virus replication. Ann N Y Acad Sci 951 207 219
8. ShiSTLaiMM 2005 Viral and cellular proteins involved in coronavirus replication. Curr Top Microbiol Immunol 287 95 131
9. PanavasTNagyPD 2003 Yeast as a model host to study replication and recombination of defective interfering RNA of Tomato bushy stunt virus. Virology 314 315 325
10. PanavieneZPanavasTServaSNagyPD 2004 Purification of the cucumber necrosis virus replicase from yeast cells: role of coexpressed viral RNA in stimulation of replicase activity. J Virol 78 8254 8263
11. PanavieneZPanavasTNagyPD 2005 Role of an internal and two 3′-terminal RNA elements in assembly of tombusvirus replicase. J Virol 79 10608 10618
12. PoganyJWhiteKANagyPD 2005 Specific Binding of Tombusvirus Replication Protein p33 to an Internal Replication Element in the Viral RNA Is Essential for Replication. J Virol 79 4859 4869
13. NagyPDPoganyJ 2008 Multiple roles of viral replication proteins in plant RNA virus replication. Methods Mol Biol 451 55 68
14. KushnerDBLindenbachBDGrdzelishviliVZNoueiryAOPaulSM 2003 Systematic, genome-wide identification of host genes affecting replication of a positive-strand RNA virus. Proc Natl Acad Sci U S A 100 15764 15769
15. PanavasTServieneEBrasherJNagyPD 2005 Yeast genome-wide screen reveals dissimilar sets of host genes affecting replication of RNA viruses. Proc Natl Acad Sci U S A 102 7326 7331
16. JiangYServieneEGalJPanavasTNagyPD 2006 Identification of essential host factors affecting tombusvirus RNA replication based on the yeast Tet promoters Hughes Collection. J Virol 80 7394 7404
17. NagyPDPoganyJ 2010 Global genomics and proteomics approaches to identify host factors as targets to induce resistance against tomato bushy stunt virus. Adv Virus Res 76 123 177
18. WangRYStorkJPoganyJNagyPD 2009 A temperature sensitive mutant of heat shock protein 70 reveals an essential role during the early steps of tombusvirus replication. Virology 394 28 38
19. WangRYStorkJNagyPD 2009 A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to intracellular membranes. J Virol 83 3276 3287
20. PoganyJStorkJLiZNagyPD 2008 In vitro assembly of the Tomato bushy stunt virus replicase requires the host Heat shock protein 70. Proc Natl Acad Sci U S A 105 19956 19961
21. ServaSNagyPD 2006 Proteomics analysis of the tombusvirus replicase: Hsp70 molecular chaperone is associated with the replicase and enhances viral RNA replication. J Virol 80 2162 2169
22. LiZBarajasDPanavasTHerbstDANagyPD 2008 Cdc34p Ubiquitin-Conjugating Enzyme Is a Component of the Tombusvirus Replicase Complex and Ubiquitinates p33 Replication Protein. J Virol 82 6911 6926
23. LiZPoganyJTupmanSEspositoAMKinzyTG 2010 Translation Elongation Factor 1A Facilitates the Assembly of the Tombusvirus Replicase and Stimulates Minus-Strand Synthesis. PLoS Pathog 6 e1001175
24. LiZPoganyJPanavasTXuKEspositoAM 2009 Translation elongation factor 1A is a component of the tombusvirus replicase complex and affects the stability of the p33 replication co-factor. Virology 385 245 260
25. PathakKBSasvariZNagyPD 2008 The host Pex19p plays a role in peroxisomal localization of tombusvirus replication proteins. Virology 379 294 305
26. BarajasDNagyPD 2010 Ubiquitination of tombusvirus p33 replication protein plays a role in virus replication and binding to the host Vps23p ESCRT protein. Virology 397 358 368
27. BarajasDJiangYNagyPD 2009 A Unique Role for the Host ESCRT Proteins in Replication of Tomato bushy stunt virus. PLoS Pathog 5 e1000705
28. MateyakMKKinzyTG 2010 eEF1A: thinking outside the ribosome. J Biol Chem 285 21209 21213
29. EspositoAMKinzyTG 2010 The eukaryotic translation elongation Factor 1Bgamma has a non-guanine nucleotide exchange factor role in protein metabolism. J Biol Chem 285 37995 38004
30. OlarewajuOOrtizPAChowdhuryWQChatterjeeIKinzyTG 2004 The translation elongation factor eEF1B plays a role in the oxidative stress response pathway. RNA Biol 1 89 94
31. DasTMathurMGuptaAKJanssenGMBanerjeeAK 1998 RNA polymerase of vesicular stomatitis virus specifically associates with translation elongation factor-1 alphabetagamma for its activity. Proc Natl Acad Sci U S A 95 1449 1454
32. Le SourdFBoulbenSLe BouffantRCormierPMoralesJ 2006 eEF1B: At the dawn of the 21st century. Biochim Biophys Acta 1759 13 31
33. KinzyTGRipmasterTLWoolfordJLJr 1994 Multiple genes encode the translation elongation factor EF-1 gamma in Saccharomyces cerevisiae. Nucleic Acids Res 22 2703 2707
34. PoganyJNagyPD 2008 Authentic replication and recombination of Tomato bushy stunt virus RNA in a cell-free extract from yeast. J Virol 82 5967 5980
35. ChengCPPanavasTLuoGNagyPD 2005 Heterologous RNA replication enhancer stimulates in vitro RNA synthesis and template-switching by the carmovirus, but not by the tombusvirus, RNA-dependent RNA polymerase: implication for modular evolution of RNA viruses. Virology 341 107 121
36. RajendranKSPoganyJNagyPD 2002 Comparison of turnip crinkle virus RNA-dependent RNA polymerase preparations expressed in Escherichia coli or derived from infected plants. J Virol 76 1707 1717
37. ChengCPNagyPD 2003 Mechanism of RNA recombination in carmo- and tombusviruses: evidence for template switching by the RNA-dependent RNA polymerase in vitro. J Virol 77 12033 12047
38. NagyPDPoganyJ 2000 Partial purification and characterization of Cucumber necrosis virus and Tomato bushy stunt virus RNA-dependent RNA polymerases: similarities and differences in template usage between tombusvirus and carmovirus RNA-dependent RNA polymerases. Virology 276 279 288
39. PoganyJFabianMRWhiteKANagyPD 2003 A replication silencer element in a plus-strand RNA virus. Embo J 22 5602 5611
40. PathakKBNagyPD 2009 Defective Interfering RNAs: Foes of Viruses and Friends of Virologists. Viruses-Basel 1 895 919
41. WhiteKANagyPD 2004 Advances in the molecular biology of tombusviruses: gene expression, genome replication, and recombination. Prog Nucleic Acid Res Mol Biol 78 187 226
42. JonczykMPathakKBSharmaMNagyPD 2007 Exploiting alternative subcellular location for replication: tombusvirus replication switches to the endoplasmic reticulum in the absence of peroxisomes. Virology 362 320 330
43. PanavasTHawkinsCMPanavieneZNagyPD 2005 The role of the p33:p33/p92 interaction domain in RNA replication and intracellular localization of p33 and p92 proteins of Cucumber necrosis tombusvirus. Virology 338 81 95
44. JohnsonCMPerezDRFrenchRMerrickWCDonisRO 2001 The NS5A protein of bovine viral diarrhoea virus interacts with the alpha subunit of translation elongation factor-1. J Gen Virol 82 2935 2943
45. QanungoKRShajiDMathurMBanerjeeAK 2004 Two RNA polymerase complexes from vesicular stomatitis virus-infected cells that carry out transcription and replication of genome RNA. Proc Natl Acad Sci U S A 101 5952 5957
46. YamajiYSakuraiKHamadaKKomatsuKOzekiJ 2010 Significance of eukaryotic translation elongation factor 1A in tobacco mosaic virus infection. Arch Virol 155 263 268
47. GavinACBoscheMKrauseRGrandiPMarziochM 2002 Functional organization of the yeast proteome by systematic analysis of protein complexes. Nature 415 141 147
48. GavinACAloyPGrandiPKrauseRBoescheM 2006 Proteome survey reveals modularity of the yeast cell machinery. Nature 440 631 636
49. CollinsSRKemmerenPZhaoXCGreenblattJFSpencerF 2007 Toward a comprehensive atlas of the physical interactome of Saccharomyces cerevisiae. Mol Cell Proteomics 6 439 450
50. PanavasTPoganyJNagyPD 2002 Analysis of minimal promoter sequences for plus-strand synthesis by the Cucumber necrosis virus RNA-dependent RNA polymerase. Virology 296 263 274
51. Al-MaghrebiMBruleHPadkinaMAllenCHolmesWM 2002 The 3′ untranslated region of human vimentin mRNA interacts with protein complexes containing eEF-1gamma and HAX-1. Nucleic Acids Res 30 5017 5028
52. BlumenthalTYoungRABrownS 1976 Function and structure in phage Qbeta RNA replicase. Association of EF-Tu-Ts with the other enzyme subunits. J Biol Chem 251 2740 2743
53. BlumenthalTLandersTAWeberK 1972 Bacteriophage Q replicase contains the protein biosynthesis elongation factors EF Tu and EF Ts. Proc Natl Acad Sci U S A 69 1313 1317
54. DreherTW 1999 Functions of the 3′-Untranslated Regions of Positive Strand Rna Viral Genomes. Annu Rev Phytopathol 37 151 174
55. ThiviergeKCottonSDufresnePJMathieuIBeaucheminC 2008 Eukaryotic elongation factor 1A interacts with Turnip mosaic virus RNA-dependent RNA polymerase and VPg-Pro in virus-induced vesicles. Virology 377 216 225
56. NishikioriMDohiKMoriMMeshiTNaitoS 2006 Membrane-bound tomato mosaic virus replication proteins participate in RNA synthesis and are associated with host proteins in a pattern distinct from those that are not membrane bound. J Virol 80 8459 8468
57. ZeenkoVVRyabovaLASpirinASRothnieHMHessD 2002 Eukaryotic elongation factor 1A interacts with the upstream pseudoknot domain in the 3′ untranslated region of tobacco mosaic virus RNA. J Virol 76 5678 5691
58. De Nova-OcampoMVillegas-SepulvedaNdel AngelRM 2002 Translation elongation factor-1alpha, La, and PTB interact with the 3′ untranslated region of dengue 4 virus RNA. Virology 295 337 347
59. BastinMHallTC 1976 Interaction of elongation factor 1 with aminoacylated brome mosaic virus and tRNA's. J Virol 20 117 122
60. SikoraDGreco-StewartVSMironPPelchatM 2009 The hepatitis delta virus RNA genome interacts with eEF1A1, p54(nrb), hnRNP-L, GAPDH and ASF/SF2. Virology 390 71 78
61. DubeABisaillonMPerreaultJP 2009 Identification of proteins from prunus persica that interact with peach latent mosaic viroid. J Virol 83 12057 12067
62. JeppesenMGOrtizPShepardWKinzyTGNyborgJ 2003 The crystal structure of the glutathione S-transferase-like domain of elongation factor 1Bgamma from Saccharomyces cerevisiae. J Biol Chem 278 47190 47198
63. NagyPDCarpenterCDSimonAE 1997 A novel 3′-end repair mechanism in an RNA virus. Proc Natl Acad Sci U S A 94 1113 1118
64. ZhangYNijbroekGSullivanMLMcCrackenAAWatkinsSC 2001 Hsp70 molecular chaperone facilitates endoplasmic reticulum-associated protein degradation of cystic fibrosis transmembrane conductance regulator in yeast. Mol Biol Cell 12 1303 1314
65. JaagHMNagyPD 2009 Silencing of Nicotiana benthamiana Xrn4p exoribonuclease promotes tombusvirus RNA accumulation and recombination. Virology 386 344 352
66. JaagHMPoganyJNagyPD 2010 A host Ca2+/Mn2+ ion pump is a factor in the emergence of viral RNA recombinants. Cell Host Microbe 7 74 81
Štítky
Hygiena a epidemiológia Infekčné lekárstvo LaboratóriumČlánok vyšiel v časopise
PLOS Pathogens
2011 Číslo 12
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Očkování proti virové hemoragické horečce Ebola experimentální vakcínou rVSVDG-ZEBOV-GP
- Koronavirus hýbe světem: Víte jak se chránit a jak postupovat v případě podezření?
Najčítanejšie v tomto čísle
- Controlling Viral Immuno-Inflammatory Lesions by Modulating Aryl Hydrocarbon Receptor Signaling
- Fungal Virulence and Development Is Regulated by Alternative Pre-mRNA 3′End Processing in
- Epstein-Barr Virus Nuclear Antigen 3C Stabilizes Gemin3 to Block p53-mediated Apoptosis
- Engineered Immunity to Infection