
Stanovení mozečkové dominance podle asymetrie svalového tonu končetin
Determination of Cerebellar Dominance from Muscle Tone of the Limbs
Objective:
Is the more pronounced physiological passivity of the left upper and lower extremities (UE, LE) in right-handed individuals equally present in the right limbs of left-handed subjects? May we define cerebellar dominance from this information?
Methods:
Muscle tone of the limbs was investigated in healthy 18–57 years old individuals (n = 69) considered right-handed (n = 26) or left-handed (n = 35) as assessed by the Edinburgh Inventory. Impaired ambidextrous (n = 8) were excluded. Muscle tone was ascertained: a) by conventional clinical examination of passivity in the shoulder, elbow, wrist, knee and instep; b) by 10 times repeated and registered number of UE swings fallowing a fall from forward arm raise, the number of shank swings following a fall from extension in sitting individuals, forearms falling from extension in prone position and after patellar and tricipital reflexes. The data were analysed using the chi-squared test of independence in 2 × 2 contingency tables; graphical presentations are based on statistical significance of percent predictive values. Number of swings was evaluated using the 2-way ANOVA model.
Results:
Right-handed females (n = 13) and right-handed males (n = 13) had a reduced muscle tone in 90% of left UEs and 65% of left LEs. Unlike right-handed, only 50% of left-handed females (n = 21) and left-handed males (n = 14) had an increased muscle tone on both extremities. The lateral difference in muscle tone, more manifested on UE, was not as evident in left-handers as in right-handers. The different pendular responses were significant in falling of forearms (p = 0.026) and in patellar reflex (p = 0.030).
Conclusions:
Right-handers represent a more homogeneous group. The asymmetry of muscle tone in left-handed does not correspond to the simple traditional concept of cerebellar dominance contralateral to the dominant hemisphere of the brain.
Key words:
muscle tone – cerebellar dominance – dextrals – left-handed – ambidextrous
Authors:
J. Tichý 1; J. Běláček 2; M. Nykl 1; N. Kaspříková 2
Authors‘ workplace:
1. LF UK a VFN v Praze
Neurologická klinika
1; 1. LF UK a VFN v Praze
Ústav biofyziky a informatiky
2
Published in:
Cesk Slov Neurol N 2012; 75/108(3): 334-343
Category:
Original Paper
Overview
Cíl:
Je fyziologicky větší pasivita na levostranných horních končetinách (HK) a na končetinách dolních (DK) srovnatelně přítomna na pravostranných končetinách u leváků? Můžeme stanovením fyziologického svalového tonu na končetinách definovat mozečkovou dominanci?
Metody:
Vyšetřovali jsme svalový tonus u zdravých osob (n = 69) ve věku 18–57 let hodnocených podle Edinburghského dotazníku jako Praváci (n = 26) a Leváci (n = 35). Rušivé ambidextry (n = 8) jsme vyřadili. Svalový tonus jsme zjišťovali: a) konvenčním klinickým vyšetřením pasivity v rameni, lokti, zápěstí, koleni a v nártu; b) desetkrát opakovaným a registrovaným počtem kyvů na HK po pádu z předpažení, na DK pak počty kyvů po pádu bérců z extenze u sedících, předloktí po pádu z extenze v lokti vleže na břiše a po vyvolání patelárních a tricipitových reflexů. Data byla zpracována standardní metodikou chí-kvadrát testu nezávislosti ve čtyřpolních tabulkách, grafická schémata jsou založena na statistické významnosti procenta prediktivních hodnot. Počty kyvů byly vyhodnoceny metodikou dvoufaktorová ANOVA.
Výsledky:
Ve skupině pravaček (n = 13) a praváků (n = 13) byl svalový tonus nižší na levé HK u 90 % vyšetřených, na levé DK u 65%. U levaček (n = 21) a u leváků (n = 14) byl na rozdíl od pravorukých vyšší svalový tonus na HK i na DK jen u 50 % vyšetřovaných! Rozdíl ve svalovém tonu, více vyjádřený na HK, nebyl u leváků tak zjevný jako u praváků. Stranově rozdílné kyvadlové odpovědi byly signifikantní při pádech předloktí (p = 0,026) a u patelárních reflexů (p = 0,030).
Závěry:
Praváci představují homogennější skupinu. U leváků asymetrie ve svalovém tonu na končetinách neodpovídá jednoduché koncepci mozečkové dominance, protilehlé hemisferální dominanci velkého mozku.
Klíčová slova:
svalový tonus – cerebelární dominance – praváci – leváci – ambidextři
Úvod
Na aktuálním stavu svalového tonu se účastní řada neuronálních okruhů propojujících nejméně těchto šest struktur: spinální míchu, mozeček, vestibulární jádra, retikulární formaci, extrapyramidový systém a kortex hemisfér [1].
Klíčovou roli představuje systém gama kličky, někdy nazývaný malý motorický systém. Zpětnovazební regulace svalového tonu, mající statickou a kinetickou složku, je dynamická funkce, jíž se účastní i struktury mozečku, u člověka včetně neocerebela. V našem dosavadním klinickém výzkumu o dominanci mozečku jsme vycházeli z tradičních výzkumů K. Hennera a jeho žáků [2–6]. Podle nich lze u praváků nalézt nižší svalový tonus na levostranných končetinách, tj. na levé ruce a na levé noze, jak odpovídá anatomickým okruhům kortiko-ponto-cerebelárních drah a zpětnovazebnímu dentato-thalamo-kortikálnímu propojení.
Zánikový neocerebelární syndrom za patologických stavů je dobře znám [7]. Zvýšená pasivita (hypotonie) na končetinách je jednou z jeho součástí. Regulace svalového tonu je velmi složitá (viz učebnice neurologie a neurofyziologie). Je otázka, zda asymetrie svalového tonu na končetinách u zdravých osob nám může určit cerebelární dominanci.
U řeči bylo prokázáno, že dominantní neocerebelum je díky kortiko-ponto-cerebelárním spojům na druhé, zkřížené straně mozečku, než jsou řečová centra velkého mozku [8–17].
Shoda zkřížené cerebro-cerebelární řečové dominance u praváků s řečovými centry v levé hemisféře a u leváků, jež mají řečová centra v pravé hemisféře, byla prokázána [12].
Aktuálním otázkám afaziologie, laterality a funkční magnetické rezonance (fMR) bylo věnováno nepřehledné množství prací, [17–42]. Umístění řečových center v levé hemisféře a „pravorukost“ nejsou zcela ve statistické shodě [18–20,23,26,27]. Většina leváků (asi tři čtvrtiny) má řečová centra v levé hemisféře jako praváci [20,36], kolem 10 % „levorukých“ má obě řečová centra a centra symbolických funkcí v pravé hemisféře, stejný počet leváků má řečová centra v obou hemisférách. Nové metodologie včetně pozitronové emisní tomografie (PET), fMR, „kognitivních evokovaných potenciálů“ (Event-Related Potentials, ERP) a přímých záznamů kortiko-subkortikálních elektrických potenciálů během neurochirurgických operací ukázaly, že klasické dělení a lokalizace řečových center je daleko složitější a variabilnější, zejména u ne-praváků [22,24,26–28]. Uložení center řeči v pravé hemisféře u praváků bylo zjištěno u 2–10 % jako tzv. zkřížená řečová dominance [33]. Pojmu „crossed dominance“, což je neodpovídání protilehlého uložení řečových center a motorické preference končetin, se v poslední době věnovala řada autorů [19,21,30,33,34,36]. Obrovský pokrok v poznávání neuronálních propojení mozku a mozečku přinesly moderní zobrazovací metody [15,16,23,25,26,36,39–44].
Mozeček je zapojen do motorických okruhů mozku a míchy. Z míchy kromě zadních provazců přivádějí proprioceptivní informace obě spinocerebelární dráhy cestami, které se liší pro horní končetiny (HK) a pro končetiny dolní (DK). Terminální informace pro Purkyňovy buňky paravermisu cerebela jsou z HK i DK homolaterální [45,46].
Spojení z dolních končetin dorzální spinocerebelární drahou (hrudní a lumbální segmenty) vychází z druhého neuronu: interneuronů v Rexedových laminách míchy (C7–L3) pro trupové svalstvo a svaly na kořeni DK. Jejich axony jako homolaterálně uložená vlákna končí nezkříženě ve vermis a paravermální oblasti mozečku [45,46]. Tractus spinocerebellaris ventralis má první neuron ve spinálních gangliích a druhý neuron ve shodných Rexedových zónách, ale pro segmenty L1–S. Velká část jejich axonů se ještě ve stejné výši míchy kříží a přikládá k fasciculus cuneatus zadních provazců. Od dráhy zadních provazců se v kmeni oddělí, kříží v horním pontu a cestou brachia conjunctiva (pedunculi cerebelli craniales) končí v homolaterální části spinocerebela. Zatímco tractus spinocerebellaris dorsalis zprostředkuje kinetickou i statickou propriocepci, ventrální spinocerebelární dráha reaguje na podněty uložené v motorické paměti jedince, přicházející „shora“ vnitřně utvořenou rytmickou informací, tj. z kortexu a z podkorových center zúčastněných na plánování a provedení hybnosti. Aktivita zpětnovazební kontroly pohybů, prokazatelná elektrofyziologicky, neustává i po přerušení zadních kořenů korespondujících segmentů [46].
Jak jsme uvedli, spinocerebelární dráhy končí homolaterálně v příslušné polovině vermis a v paravermálních oblastech kůry spinálního mozečku. Zpětnovazební regulace z paravermálních Purkyňových buněk probíhá do druhostranného nucl. ruber a odtud pokračuje zkříženou rubrospinální drahou ke kořenovému svalstvu končetin (zejména extenzorů DK). Nucleus interpositus spinocerebela hraje klíčovou roli v časování pohybů. Jeho porucha disfacilitací (inhibicí) rubrospinálních a kortikospinálních drah vede k cerebelární hypotonii a k vzniku pendulárních reflexů [46,54], nejspíše chyběním útlumu antagonistů.
Spino-cerebelární propriocepce z horních končetin ze segmentů C1–Th5 probíhá separátní drahou (tractus spino-cuneo-cerebellaris) homolaterálně v laterálních zadních provazcích do jádra nucl. cuneatus lateralis a odtud axony tohoto třetího neuronu probíhají homolaterálně v corpus restiforme do spinálního cerebela jako mechová vlákna pro Purkyňovy buňky. Spino-cuneo-cerebelární dráha zprostředkuje zejména kinetickou část hluboké citlivosti, čímž se liší od předchozích tractus spinocerebellares [45,46].
Neocerebelum (cerebro-cerebelum) kromě zpětnovazební kontroly hybnosti zabezpečuje plánování motorického jednání na podkladě aktuální percepce a na kognitivních funkcích, uložených v paměti. Mozeček má i další neurokognitivní funkce, např. rozlišení délky trvání tónu či rychlosti pohybujících se předmětů a časování [47]. Mozeček představuje v mozku neuronální počítač [52,53].
V této práci jsme se nezabývali ani časováním, ani kognitivními funkcemi mozečku včetně funkcí řeči.
Soubor a metodika
Vyšetřili jsme 69 „zdravých“ dospělých osob (muži/ženy = 33/36) ve věku 18–57 let, u kterých stupeň pravo-levorukosti byl stanoven podle Edinburghského dotazníku [48]. Svalový tonus na horních končetinách (HK) jsme klinicky hodnotili aspexí, pohmatem a pozorováním rozsahu pohybu v kloubech ramenních, v zápěstí a v lokti. Podobný postup jsme užili při vyšetřování pasivity na DK. Podrobnější postup při sledování svalového tonu bylo zaznamenání počtu kyvů paže při modifikované zkoušce „ressaut“, kdy stojící osoba uvolní rychle HK z předpažení. V poloze vleže na břiše jsme hodnotili počty kyvů po současném uvolnění HK z exteze v loketních kloubech a po vyvolání tricipitových reflexů. Dále jsme hodnotili počty kyvů u pádu bérců po současném rychlém uvolnění DK z extenze vsedě a po patelárním reflexu. Všechna jednotlivá pozorování jsme prováděli desetkrát.
Jiný postup jsme volili pro výzkumu pasivity končetin pozorováním a zaznamenáním počtu kyvů paže při modifikované zkoušce „ressaut“, kdy stojící osoba uvolní rychle HK z předpažení. V poloze vleže na břiše jsme hodnotili počty kyvů po současném uvolnění HK z exteze v loketních kloubech a po vyvolání tricipitových reflexů. Dále jsme hodnotili počty kyvů u pádu bérců po současném rychlém uvolnění DK z extenze vsedě a po patelárním reflexu vsedě. Všechna jednotlivá pozorování jsme opakovali desetkrát.
Klasifikace probandů do laterálních skupin byla v tomto případě provedena na základě „měr pravé laterality (MPL)“ definovaných (viz také v [6]) individuálním skórem ‘0’ (při „odpovědi“ vlevo), ‘1’ (při “odpovědi” vpravo) nebo ‘0,5’ (nerozlišitelná preference obou končetin) pro každou z 10 standardních otázek E-dotazníku. Za „Leváky“ byli označeni probandi, pro které byl součet všech deseti MPL shledán v rozmezí hodnot ‘0–2,5‘ a za „Praváky“ ti, pro které vyšel součet MPL na všech deseti otázkách v rozmezí ‘7,5–10‘. (Kriteriální hodnoty ‘2,5’ a ‘7,5’ byly vytipovány a prověřeny jako optimální z hlediska nejvyšší senzitivity pro všechny statistické testy, které jsme shledali jako relevantní pro všechny typy měřených indikátorů. Výše popsaným způsobem byli probandi přerozděleni do skupin: 21 Levaček a 14 Leváků (n = 35); 13 Pravaček a 13 Praváků (n = 26), resp. 8 „Ambidextrů“ (2 muži a 6 žen). Nejednoznačně se projevující skupina Ambidextrů byla posléze „jako rušivá“ vyloučena z analýz. Naše výsledky dobře korespondují s výsledky Oldfielda [48].
Vyšetření pasivity u probandů hodnocených podle deseti položek E-dotazníku bylozpracováno standardní metodikou χ2-testu nezávislosti ve čtyřpolních tabulkách. Následně byla odhadnuta „procenta prediktivních hodnot“ (marginální pravděpodobnosti souhlasné klasifikace vpravo, resp. vlevo) a na základě standardních výstupních sestav analýzy ROC [51] byla vyčíslena jejich statistická významnost vůči referenční hodnotě 50 %. Statisticky nejvýznamnější vazby mezi klinickým ohodnocením větší pasivity v pěti kloubních lokalitách, odpověďmi na deset otázek E-dotazníku a laterálními skupinami byly promítnuty do grafického schématu, které jsme porovnali s metodicky stejně zpracovaným schématem pro soubor školáků popsaný a vyšetřený v [5–6].
Počty kyvů (převzatých jako průměry z 10 opakovaných měření při samovolných pádech končetin) byly vyhodnoceny metodikou dvoufaktorové ANOVA s jedním faktorem fixním (Praváci-Leváci) a jedním vnitřním faktorem (pro „opakovaná měření“ na levé a na pravé straně HK nebo DK). Křížově dominantní pasivita, tj. nižší svalový tonus u Praváků vlevo a u Leváků vpravo, se měla projevit na úrovni tzv. interakčních efektů, které posuzují statisticky významné rozdíly v počtech kyvů (u Praváků vs Leváků) měřených ‘vlevo‘ a ‚vpravo‘ na HK nebo DK. Statisticky nejvýznamnější výsledky jsou prezentovány v grafech vytvořených v SW Statistica (verze 9). Analogický ANOVA model (pro Praváky vs Nepraváky) byl použit rovněž při prezentacích [49,50].
Výsledky
U 13 Pravaček (průměrný věk = 29,5) a 13 Praváků (průměrný věk = 30,9) byla klinicky zjištěna větší pasivita (hypotonie) na levé horní končetině shodně u obou pohlaví (graf 1a). Na levostranných DK se větší pasivita (hypotonie) u pravorukých žen i mužů vyskytovaly u 65 % (graf 1b). Bez ohledu na pohlaví se podíly větší pasivity u praváků vůči levákům lišily se spolehlivostí 95 % u zápěstí, nelišily na dolních končetinách (koleno a kotník). Ženy měly signifikantně vyšší pasivitu na lokti a u kotníku. Statistická významnost diferencí mezi skupinami je v těchto případech ovšem značně závislá na počtech probandů, z nichž jsou procenta zobrazená na grafu 1b vypočtena.


Průměrné počty kyvů zjištěné v laterálních skupinách (Praváci-Leváci) při jednotlivých zkouškách na levých (sin) a pravých (dx) HK a DK jsou uvedeny v tab. 1.
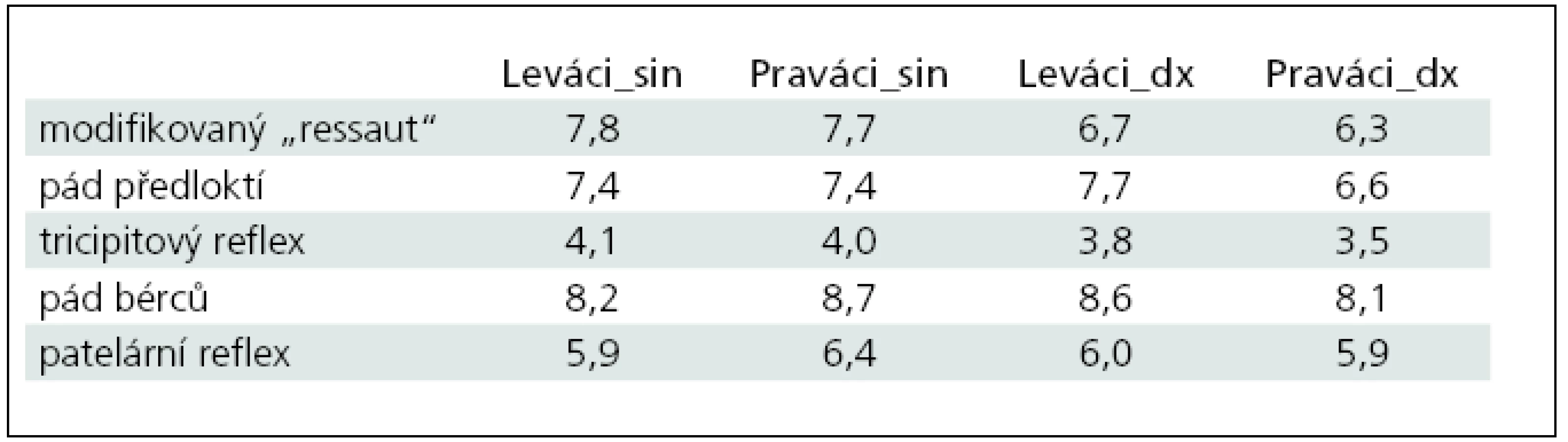
Nejvyšší počty kyvů (více než 8,0) jsme zaznamenali u volného pádu bérců, dále u pádu předloktí (s výjimkou nálezů dx u Praváků ≥ 7,4) a při volném pádu HK z předpažení u nálezů sin. Počty kyvů po reakci na poklep neurologickým kladívkem byly nejnižší (nejméně transparentní) u tricipitového reflexu (na úrovni uváděných průměrů ≤ 4,1), u patelárního reflexu v průměru kolem šesti kyvů.
Grafické vyjádření včetně posouzení statistické významnosti rozdílů námi uváděných hodnot v rámci jednotlivých zkoušek je prezentováno na grafech 2a–e. Při souběžném pádu HK z předpažení (modifikovaný „ressaut“) se nálezy ‘sin’ a ‘dx‘ v průměrných počtech kyvů liší vysoce statisticky významně (F[1;57] = 51,995; p < 0,001), nikoli však ve skupinách Praváků vůči Levákům (graf 2a). Stručně řečeno: větší pasivita v levém rameni se u Leváků neliší od Praváků!
![Graf 2a. Porovnání průměrného počtu kyvů (z 10 opakování pokusu) při současném samovolném pádu paží z předpažení ve stoji (modifikovaný „ressaut“) podle lokality (sin-dx) a laterální skupiny (Praváci-Leváci).
Elipsy propojují statisticky významně nerozlišitelné průměry (pro Praváky nebo Leváky v týchž lokalitách). Nálezy poukazují na statisticky významně nižší svalový tonus na levých HK, efekt křížové dominance HK (testovaný interakčním efektem ‘sin-dx‘ a laterální skupiny) nebyl shledán jako signifikantní (F[1,57] = 0,777; p = 0,382).](https://pl-master.mdcdn.cz/media/image/a871aeb70a038acb08dd5106f3298902.png?version=1537795798)
Při souběžném pádu předloktí vleže se ale křížově dominantní končetiny průměrným počtem kyvů liší (F[1,54] = 5,207; p = 0,026 – graf 2b). Právě tak se liší i nálezy sin vůči dx u Praváků (pro odpovídající post-hoc test; p = 0,035), u Leváků však již nikoli statisticky významně (p = 0,309). Po reakci na provedení tricipitového reflexu vleže se nám však (podobně jako u modifikovaného „ressaut“) křížovou dominanci Praváků vůči Levákům na HK nepodařilo verifikovat (graf 2c). Rovněž v tomto případě se ale zjištěné průměry počtu kyvů lišily statisticky významně pro sin vůči dx (F[1,57] = 18,095; p < 0,001), a to i odděleně ve skupině Praváků (pro post-hoc test; p < 0,001) i ve skupině Leváků (p = 0,030).
![Graf 2b. Porovnání průměrného počtu kyvů (z 10 opakování pokusu) při současném pádu předloktí z extenze vleže podle lokality (sin-dx) a laterální skupiny (Praváci-Leváci).
Hyperboly poukazují na statisticky významně rozlišitelné průměry u Praváků (na rozdíl od Leváků) na hl. spolehlivosti 95 %. Efekt křížové dominance HK (testovaný interakčním efektem ‘sin-dx‘ a laterální skupiny) je statisticky významný na hladině spolehlivosti
95% (F[1,54] = 5,207; p = 0,026).](https://pl-master.mdcdn.cz/media/image/69da3f16ea0195d3590a7db466e6097f.png?version=1537794431)
![Graf 2c. Porovnání průměrného počtu kyvů (z 10 opakování pokusu) po reakci tricipitového reflexu podle lokality (sin-dx) a laterální skupiny (Praváci-Leváci).
Elipsy propojují statisticky významně nerozlišitelné průměry (pro Praváky nebo Leváky v týchž lokalitách). Nálezy poukazují na statisticky významně nižší svalový tonus na LHK, efekt křížové dominance HK (testovaný interakčním efektem ‘sin-dx‘ a laterální skupiny) je nesignifikantní (F[1,57] = 1,857; p = 0,178).](https://pl-master.mdcdn.cz/media/image/b8915b06e31b0038462bae0ea7cb79c3.png?version=1537796187)
Průměrné počty kyvů u současného pádu bérců z extenze vsedě křížovou dominanci laterality vůči svalovému tonu potvrzují (F[1,54] = 5,934; p = 0,018), a to v principu symetricky u obou DK (graf 2d). DK měřené počtem kyvů po reakci na provedený patelární reflex byly rovněž shledány jako křížově dominantní (F[1,55] = 4,962; p = 0,030), právě tak jako rozdíly sin vůči dx ve skupině Praváků (pro post-hoc test; p = 0,018) (graf 2e). Na úrovni DK jsou tedy naše výsledky konzistentnější s předpokladem křížové dominance rukosti vůči registraci nižšího svalového tonu na protilehlých končetinách. Dost možná je to způsobeno i tím, že u těchto klinických zkoušek je evidence počtu kyvů nejzjevnější.
![Graf 2d. Porovnání průměrného počtu kyvů (z 10 opakování pokusu) při současném pádu bérců z extenze vsedě podle lokality (sin-dx) a laterální skupiny (Praváci-Leváci).
Elipsa indikuje statisticky významně nerozlišitelné průměry. Efekt křížové dominance DK (testovaný interakčním efektem ‘sin-dx‘ a laterální skupiny) je statisticky významný na hladině spolehlivosti 95% (F[1,54] = 5,934; p = 0,018).](https://pl-master.mdcdn.cz/media/image/179606cfc4986d90143bb67fbb1dfa34.png?version=1537792638)
![Graf 2e. Porovnání průměrného počtu kyvů (z 10 opakování pokusu) po reakci patelárního reflexu podle lokality (sin-dx) a laterální skupiny (Praváci-Leváci).
Hyperboly poukazují na statisticky významně rozlišitelné průměry u Praváků (na rozdíl od Leváků) na hladině spolehlivosti 95 %. Efekt křížové dominance DK (testovaný interakčním efektem ‘sin-dx‘ a laterální skupiny) je statisticky významný na hladině spolehlivosti 95% (F[1,55] = 4,96; p = 0,030).](https://pl-master.mdcdn.cz/media/image/23ba6c22cabc0d0af0d91c856b44f769.png?version=1537792992)
Diskuze
Problém laterality zajímá laickou i odbornou veřejnost velmi dlouho. Za klinicky nejzávažnější problematiku lze považovat umístění řečových center v tzv. dominantní mozkové hemisféře. Proto jsme se v úvodu podrobněji zmínili o lokalizaci řeči v mozku a v mozečku. Díky moderním zobrazovacím metodám a velkému počtu prací na zdravých jedincích víme, že kolem 90–95 % jedinců ve všech populacích „hovoří“ levou hemisférou. Díky anatomům jsou dobře dokumentována zkřížená spojení kortiko-ponto-cerebelární. Mozečková funkční asymetrie cerebro-cerebelárními okruhy je velice složitá [15,16]. Pro relativně malé počty probandů a zkušenost, že Leváci, resp. Ambidextři tvoří méně homogenní skupinu, lze při výzkumu laterality výsledky posuzovat též v uspořádání sestavy rozdělené na Praváky a Nepraváky. V rámci této studie použitá klasifikace do laterálních skupin vymezuje Leváky na bázi jednotného (více symetrického) metodického konceptu. To umožňuje objektivnější diskriminaci Praváků vůči Levákům a zvyšuje sílu statistických testů pro většinu námi použitých laterálních ukazatelů (i z důvodu snížení počtu probandů spadajících do zbytkové skupiny Ambidextrů).
Při sledování dospělých zdravých osob jsme u více než u poloviny levorukých probandů (žen i mužů) nalezli, obdobně jako u 9–11letých dětí, nižší svalový tonus a větší pasivitu na levostranných končetinách, tedy stejně jako u praváků [5,6]. U hodnocených dětí stoupalo procento preference levé dolní končetiny od prostého přeskoku do dálky z 50 % přes poskoky po jedné, kopání do míče až po napsání písmena či číslice patou v 98 % shodně s preferencí ruky. Rukost a umístění řečových center v mozku jsme v této práci neřešili [Peterová V, Tichý J, Běláček J, Nykl M et al. Řečová hemisferální centra a mozečková dominance (in preparation)].
Pro účely systematického srovnání obou souborů (školáků vs klinicky vyšetřených dospělých) jsme provedli strukturální analýzu založenou na systematické statistické analýze prediktivních hodnot mezi indikátory pasivity v pěti lokalitách a deseti otázkami E-dotazníku (schémata 1, 2). Procenta prediktivních hodnot (vyjádřená na schématech v různých barevných odstínech podle statistické významnosti před koncovými šipkami) reprezentují pravděpodobnosti, s jakou vyšetření v lokalitě, resp. odpovědi z E-dotazníku predikují hodnotu indikátoru, do něhož směřuje šipka (záporné hodnoty poukazují na inverzní vztah pasivity k dominanci, tj. když vyšetření/odpověď ‘sin’ u jedné proměnné implikuje vyšetření/odpověď ‘dx’ u druhé proměnné a naopak); statisticky nevýznamné prediktivní hodnoty (vůči referenční hladině 50 %) se ve schématech nezobrazují.


Schémata poukazují zejména na kvalitativně významnější vztahy pasivity ramene k lokti a ke koleni u dospělých; u dětí souhyby HK při chůzi nebyly ve statistické shodě s dalšími lokalitami. Větší signifikace procenta prediktivních hodnot pro soubor školáků souvisí zjevně s více než pětinásobně větším rozsahem tohoto souboru oproti souboru dospělých. Parciálně se do odlišnosti výsledků pro oba soubory promítá i fakt, že soubor školáků byl pořízen jako reprezentativní pro 9–11leté děti pražských škol (akcentující dominantní podíl Praváků ve vzorku v poměru přibližně 9 : 1 vůči Levákům), zatímco soubor dospělých byl vytvořen selektivně s cílem postavit proti sobě dvě početně srovnatelné skupiny (ve snaze párovat Praváky vůči Levákům v poměru 1 : 1).
Hennerova škola vycházela z klinických pozorování, že praváci mají jako dominantní pravou polovinu mozečku. Fyziologickým projevem mozečkové dominance by měl být nižší svalový tonus a větší pasivita a kloubní hra na „nedominantních končetinách“. U zdravých 9–11letých školáků jsme to mohli potvrdit jen částečně. Zkřížená dominance cerebro-cerebelární platí pro Pravačky a pro Praváky dětského i dospělého věku pro horní končetinu, méně pro končetinu dolní.
Jsou pasivita a hypotonie končetin spolehlivým příznakem cerebelární dominance? Neplatí tento vztah více pro horní končetiny (HK), které jsou více spojeny s volní aktivitou, než končetiny dolní (DK), sloužící k lokomoci, převážně mimovolní? (Horní končetiny jsou těsněji svázány s cerebrálními centry symbolických funkcí – psaní, kreslení.)
U stoprocentních leváků (podle šesti navzájem nerozlišitelných otázek E-dotazníku) jsme nalezli překvapivě vysoké procento relativní hypotonie a větší pasivity podle klinického hodnocení na levostranných končetinách, tj. jako u praváků. Procentuální zastoupení pasivity u leváků bylo u nich téměř shodné na HK i na DK. Ambidextři se podle stanovení pasivity více podobali Levákům než Pravákům, pro rušivý efekt vyhodnocení dvoufaktorovou ANOVA jsme osm Ambidextrů ze souboru dospělých vyřadili.
Stupeň rukosti podle Edinburgského dotazníku a řečovou cerebrální lokalizaci sledovali Isaacs et al [29,35]. Atypická lateralita se vyskytovala u Ambidextrů a u Leváků daleko častěji než u Praváků. Podobný vztah jsme nalezli i my při sledování mozečkové dominance podle tradičního vyšetřování asymetrie svalového tonu končetin.
Míra laterality podle preference jedné z HK či DK může být testována nekonečně velkým počtem úkolů, což vede ještě k větší variabilitě v odpovědích. Vliv okolí a tlak „pravoruké civilizace“ je dostatečně znám. U starších jedinců díky přeučování ve škole je tomu tak zejména při psaní. Skupina zdravých osob ukázala, že v patřičné poloze při dokonalé relaxaci vsedě počty kyvů patelárních reflexů přesahují často hodnoty 7–8 kyvů, což je hraniční počet uváděný pro kyvadlové reflexy [54–61].
Validace metodiky měření pasivity založené na vizuálním odhadu počtu kyvů byla provedena u zkoušky „modifikovaný ressaut“ metodikou ANOVA pro opakovaná měření. Při této analýze nebyly zjištěny systematické rozdíly v rámci 10 opakování pokusu (F[9; 504]= 0,883; p = 0,540). Ke stejnému závěru autoři článku dospěli při formální analýze elektronických záznamů akcelerace spontánních kyvů u zkoušky „pád bérců z extenze“ [50]. Elektrofyziologická vyšetření elektromagnetickými čidly, umístěnými na zápěstí a nad kotníky DK, ve srovnání se zde uváděnými tradičními klinickými testy sdělíme na jiném místě.
Při klinickém stanovení rukosti a jejího stupně aspexí a palpací jsme zjistili stranu větší pasivity v kloubech ramene, lokte, zápěstí, kolene a nártu; kromě tohoto klasického neurologického posuzování včetně asymetrií synkinez HK při chůzi jsme zaznamenávali počet kyvů po pádu paží z předpažení (modifikovaná zkouška „ressaut“), počty kyvů po patelárním a tricipitovém reflexu a po zkoušce současného pádu bérců z extenze v koleni vsedě či předloktí z extenze v lokti vleže (na břiše). V našem relativně nevelikém souboru jsme vyšetřovali i řadu úkonů, které zde nejsou uváděny. Zjistili jsme, že i tradiční klinické vyšetření je schopné přinést nové informace, které po objektivizaci přístrojovým zařízením mohou rozšířit naše vědomosti o celém mysteriu laterality.
Ohodnocení větší či menší pasivity vlevo či vpravo na HK a DK palpací a aspeksí (standardní neurologický postup) jsme použili v rámci vyšetřování rukosti a nohosti v souboru 9–11letých školáků s postupným navýšením rozsahu souboru (až na n = 366) v letech 2007–2009 [5,6]. Vyhodnocení indikátorů pasivity navzájem a vůči rukosti prostřednictvím grafické prezentace statisticky významných „prediktivních hodnot“ bylo prezentováno v [49], když klasifikace do laterálních skupin byla provedena na bázi šesti testů E-dotazníku nejvýznamnějších pro rukost.
Cennou informaci o svalovém tonu přináší pozorování počtu kyvů po volném pádu bérce z extenze vsedě podle Wartenberga [54]. Jako „normální“ kyvadlové reflexy patelární se uvádějí 2–3 kyvy [54–61]. Za patologické je považován větší počet kyvů než 7–8. Podle našich zkušeností u zdravých dětí [5,6] a u zdravých dospělých je rozdíl v počtu kyvů na levostranných či pravostranných končetinách v rámci jejich tonusové asymetrie v hodnotách 1–3 kyvů. Absolutní počet kyvů je interindividuálně velice variabilní, závisí i na svalové hmotě. Muži a muskulární typy mají menší počet kyvů. Důležitá je spolupráce s vyšetřovaným, aby nedocházelo k volnímu zastavování kyvů či k jejich arteficiálnímu prodlužování, což lze zjistit pozorováním a patřičný pokus vyřadit. Artefakt lze prokázat graficky [49,50]. Útlum kyvadlových reflexů nastává u spasticity a u poruch extrapyramidových. U zánikových lézí mozečku díky hypotonii jejich počtu přibývá.
K pochopení složitosti svalového tonu obecně: člověk jako dipéd má díky gravitaci specifická spojení, ovlivňující odlišně HK a DK.
Do složitých mechanizmů regulace svalového tonu jak statického, tak kinetického jsou zapojeny okruhy kortikální: emocionální, zrakové, sluchové, subkortikální (vestibulární) a složka pohybové paměti, tedy nácviku, souvisejícího s časováním [38]. Problematice časování se u nás věnovala brněnská škola [63].
Rychlost vedení nervovými vlákny závisí na jejich anatomické skladbě a na stupni myelinizace [64].
Nenalezli jsme údaje o tom, o kolik rychleji probíhají impulzy v kortiko-ponto--cerebelárních drahách při úvaze, že od úmyslu provést jakýkoliv pohyb musí motorická kůra a přední rohy míšní obdržet předem informace o poloze těla i jeho končetin a paměťových motorických okruhů z mozečku a z mozku.
Výsledky klinického stanovení svalového tonu jsou natolik přesvědčivé, že mozečková dominance u Leváků není prostým „zrcadlovým obrazem“ cerebelární dominance Pravaček a Praváků. Ambidextři se chovají jako neúplní Leváci.
Zkřížená motorická dominance cerebro-cerebelární v Hennerově pojetí platí pravděpodobně jen pro čisté praváky a více pro jejich horní končetiny. U Praváků je nižší svalový tonus na levostranných horních i dolních končetinách u dětí i u dospělých. Ve vysokém procentu jej ale nalézáme i u Leváků a u Ambidextrů.
Charakteristiky pádů při modifikovaném „ressaut“ souhlasily s klinickým pozorováním souhybů HK při chůzi a s manuálním posouzením pasivity v ramenním kloubu. Souhyby paží při chůzi jsou někdy více vyjádřeny v loketním kloubu než v samotném rameni, což by stálo za zvláštní sledování např. i s měřením délky kroku druhostranné dolní končetiny.
Při posuzování motorické laterality lze používat dělení na Praváky a Nepraváky. Vždy musíme mít povědomí o variabilitě jednotlivých izolovaných úkonů. Znovu je třeba zdůraznit, že mozeček kromě regulace motorických funkcí se účastní na řadě kognitivních úkonů [52,63].
prof. MUDr. Jiří Tichý, CSc.
Neurologická klinika
1. LF UK a VFN
Kateřinská 30
120 00 Praha 2
e-mail: jiri.tichy@lf1.cuni.cz
Přijato k recenzi: 6. 6. 2011
Přijato do tisku: 13. 12. 2011
Sources
1. Krkljes M. Regulation of muscle Tonus. Med Pregl 1990; 43(3–4): 172–174.
2. Henner K. Příznaky vznikající z nadměrné činnosti mozečku. Praha: Grégr a syn 1928.
3. Hrbek J. Nový výklad převládající pravorukosti, dominance levé mozkové hemisféry a pravé mozečkové hemisféry. Cesk Slov Neurol N 1978; 41/74(3): 145–165.
4. Tichý J. Vír vlasů – geneticky vázaný jev při vyšetřování fenotypu praváctví a hemisferální dominance. Čas Lék čes 2006; 145 : 995–998.
5. Tichý J. Běláček J. Pravo/levorukost a preference druhostranné dolní končetiny. Testování laterality a mozečkové dominance. Cesk Slov Neurol N 2008; 71/104(5): 552–558.
6. Tichý J. Běláček J. Laterality in children: cerebellar dominance, handedness, footedness and hair whorl. Activitas Nervosa Superior Rediviva 2009; 51(1–2): 9–20.
7. Růžička E, Amber Z. Mozeček. In: Amber Z, Bednařík J, Růžička E (eds). Klinická neurologie. I. část obecná. Praha: Triton 2004.
8. Marien P, Engelborghs S, Fabbro F, De Devn PP. The lateralized linguistic cerebellum: a review and a new hypothesis. Brain Lang 2001; 79(3): 580–600.
9. Gebhart AL, Petersen SE, Thach WT. Role of the Posterolateral Cerebellum in Language. Ann N Y Acad Sci 2002; 978 : 318–333.
10. Urban PP, Marx J, Hunsche S, Gawehn J, Vucurevic G, Wicht S et al. Cerebellar Speech Representation. Arch Neurol 2003; 60(7): 965–972.
11. Hubrich-Ungureanu P, Kaemmerer N, Henn FA, Braus DF. Lateralized organization of the cerebellum in a silent verbal fluency task: a functional magnetic rezonance imaging study in healthy volunteers. Neurosci Lett 2002; 319(2): 91–94.
12. Jansen A, Flöel A, Van Randenborgh J, Konrad C, Rotte M, Förster AF et al. Crossed cerebro-cerebellar language dominance. Hum Brain Mapp 2005; 24(3): 165–172.
12. Ito M. Cerebellar circuitry as a neuronal machine. Prog Neurobiol2006; 78(3–5): 272–303.
14. De Smet HJ, Baillieux H, De Deyn PP, Mariën P, Paquier P. The cerebellum and language: the story so far. Folia Phoniatr Logop 2007; 59(4): 165–170.
15. Kajimoto K, Oku N, Kymura Y, Kato H, Tahaka MR, Kanai Y et al. Crossed cerebellar diaschisis: a positron emission tomography study. Ann Nucl Med 2007; 21(2): 109–113.
16. Krienen FM, Buckner RL. Segregated fronto-cerebellar circuits revealed by intrinsic functional connectivity. Cereb Cortex 2009; 19(10): 2485–2497.
17. Murdoch BE. The cerebellum and language: historical perspective and review. Cortex 2010; 49(7): 858–868.
18. Neff D. Differences in the function of the two cerebral hemispheres. In: Geschwind N, Galaburda AM (eds). Cerebral dominance: The biological foundations. Cambridge MA: Harvard University Press 1984.
19. Bakar M, Kirschner HS, Hertz RT. Crossed aphasia. Functional braion imaging with PET or SPECT. Arch Neurol 1996; 53(10): 1026–1032.
20. Annett M, Alexander MP. Atypical cerebral dominance: Prediction and tests of the right shift theory. Neuropsychologia 1996; 34(12): 1215–1227.
21. Chee MW, Buckner RL, Savoy RL. Right hemisphere language in a neurologically normal dextral: a fMRI study. Neuroreport 1998; 9(15): 3499–3502.
22. McManus IC. Handedness, cerebral lateralisation, and the evolution of language. In: Corballis MC, Lee SEG (eds). The descent of mind. Oxford: Oxford University Press 1999.
23. Tzourio N, Crivello F, Mellet E, Nkanga-Ngila B, Mazoyer B. Functional anatomy of dominance for speech comprehension in left handers vs right handers. Neuroimage 1998; 8(1): 1–16.
24. Annett M. Handedness and cerebral dominance: the right shift theory. J Neuropsychiatry Clin Neurosci 1998; 10(4): 459–469.
25. Pujol J, Deus J, Posilka JM, Capdevila A. Cerebral lateralization of language in normal left-handed people studied by functional MRI. Neurology 1999; 52(5): 1038–1048.
26. Knecht S, Dräger B, Deppe M, Bobe L, Lohmann A, Flöel E et al. Handedness and hemispheric language dominance in healthy humans. Brain 2000; 123(12): 2512–2518.
27. Knecht S, Deppe M, Dräger B, Bobe L, Lohmann H, Ringelstein B et al. Handedness and hemispheric language dominance in healthy humans. Brain 2000; 123(12): 2512–2518.
28. Tzourio-Mazoyer N, Josse G, Crivello F, Mazoyer B. Interindividual variability in the hemispheric organization for speech. Neuroimage 2000; 21(1): 422–435.
29. Isaacs KL, Khedr EM, Hamed E, Said A, Basahi J. Handedness and language cerebral lateralization. Eur J Appl Physiol 2002; 87(4–5): 469–473.
30. Chee MW, Caplan D. Face encoding and psychometric testing in healthy dextrals with right hemisphere language. Neurology 2002; 59(12): 1928–1934.
31. Borbély K, Gjedde A, Nyáry I, Czirják S, Donauer N, Buck A. Speech activation of language dominant hemisphere: a single-photon emission computed tomography study. Neuroimage 2003; 20(2): 987–994.
32. Wood AG, Harvey AS, Wellard RM, Abbot DF, Anderson V, Kean M et al. Language cortex activation in normal children. Neurology 2004; 63(6): 1035–1044.
33. Klateb A, Martyry MD, Annoni JM, Lazeyras F, de Tribolet N, Pegma AJ et al. Transient crossed aphasia evidence by functional brain imagery. Neuroreport 2004; 15(5): 785–790.
34. Lindelll AK. In your right mind: right hemisphere contributions to language processing and production. Neuropsychol Rev 2006; 16(3): 131–148.
35. Barr WB, Nelson PK, Devinski O. Degree of handedness and cerebral dominance. Neurology 2006; 66(12): 1855–1858.
36. Crosson B, McGregor K, Kaudinya SG, Conway TW, Benjamin M, Yu-Ling Chan et al. Functional MRI of language in aphasia: a review of the literature and the methodological challenges. Neuropsychol Rev 2007; 17(2): 157–177.
37. Hatta T. Handedness and the brain: a review of brain-imaging techniques. Magn Reson Med Sci 2007; 6(2) 99–112.
38. Keele SW, Ivry R. Does the cerebellum provide a common computation for diverse tasks? A timing hypothesis. Ann N Y Acad Sci 1990; 608 : 179–211.
39. Marien P, Engelborghs S, Fabbro F, De Devn PP. The lateralized linguistic cerebellum: a review and a new hypothesis. Brain Lang 2001; 79(3): 580–600.
40. Gebhart AL, Petersen SE, Tlach WT. Role of the posterolateral cerebellum in language. Ann N Y Acad Sci 2002; 978 : 318–333.
41. Hubrich-Ungueranu P, Kaemmerer N, Henn FA, Braus DF. Lateralized organization of the cerebellum in a silent verbal fluency task: a functional magnetic rezonance imaging study in healthy volunteers. Neurosci Lett 2002; 319(2): 91–94.
42. Urban PP, Marx J, Hunsche S, Gawehn J, Vucurevic G, Wicht S et al. Cerebellar speech representation: lesion topography in dysarthria as derived from cerebellar ischemia and functional magnetic resonance imaging. Arch Neurol 2003; 60(7): 965–972.
43. Habas C, Axelrad H, Cabanis EA. The cerebellar second homunkulus remains silent during passive bimanual movements. Neuroreport 2004; 15(10): 1571–1574.
44. Kajimoto K, Oku N, Kymura Y, Kato H, Tahaka MR, Kanai Y et al. Crossed cerebellar diaschisis: a positron emission tomography study. Ann Nucl Med 2007; 21(2): 109–113.
45. Chusid JG. Correlative Neuroanatomy and Functional Neurology. 18th ed. Los Altos, California: Lange Medical Publication 1982.
46. Kandel ER, Schwarz JH, Jessell TM. Principles of neural science. 4th ed. New York: McGraw Hill 2000.
47. Ivry RB, Keele SW. Timing functions of the cerebellum. J Cogn Neurosci 1989; 1 : 136–152.
48. Oldfield RC. The Assessment and Analysis of Handedness: The Edingurgh Inventory. Neuropsychologia 1971; 9(1): 97–113.
49. Běláček J, Tichý J, Novák M, Nykl M, Pecha O, Voleman M. Měření laterality mozečku – metodika. Poster. In: Mornstein V, Vlk D (eds). Sborník abstrakt XXXIII. dnů lékařské biofyziky Mikulov 2010.
50. Běláček J, Tichý J, Novák M, Nykl M, Pecha O, Kaspříková N. Měření laterality – statistická analýza klinických vyšetření dominance mozečku. Cesk Slov Neurol N 2010; 73/103 (Suppl 2): S113–S114.
51. SPSS Statistisc 17.0 Command Syntax Reference. Chicago, IL 60606-6412 : 1717–1720.
52. Ito M. Historical review of the significance of the cerebellum and the cells in motor learning. Ann N Y Acad Sci 2002; 978 : 273–288.
53. Ito M. Cerebellar circuitry as a neuronal machine. Prog Neurobiol 2006; 78(3–5): 272–303.
54. Wartenberg R. Pendulousness of the legs as a diagnostic test. Neurology 1951; 1(1): 18–24.
55. Vítek J. Obecná neurologická diagnostika. Praha: Zdravotnické nakladatelství 1950.
56. Černáček J et al. Neurologická propedeutika. Bratislava: Vydavatelstvo Slovenskej Akademie Vied 1963.
57. Brown RA, Lawson DA, Leslie GC, Part NJ. Observation on the applicability of the Wartenberg pendulum test to healthy, elderly subjects. J Neurol Neurosurg Psychiatry 1988; 51(9): 1171–1177.
58. Lin CC, Ju MS, Lin CW. The pendulum test evaluating spasticity of the elbow joint. Arch Phys Med Rehabil 2003; 84(1): 69–74.
59. Huang HW, Ju MS, Wang WC, Li CC. Muscle tone of upper limbs evaluated by quantitative pendulum test in patients with acute cerebral stroke. Acta Neurol Taiwan 2009; 18(4): 250–254.
60. Bohannon RW, Harrison S, Kinsella-Shaw J. Reliability and validity of pendulum test measures of spasticity obtained with the Polhemus tracking system from patients with chronic stroke. J Neuroeng Rehabil 2009; 6 : 30.
61. Růžička E, Ambler Z. Mozeček. In: Ambler Z, Bednařík J, Růžička E (eds). Klinická neurologie. 1. část obecná. Praha: Triton 2004.
62. Bares M, Lungu OV, Liu T, Waechter T, Gomez CM, Ashe J. The neural substrate of predictive motor timing in spinocerebellar ataxia. Cerebellum 2011; 10(2): 233–244.
63. Stoodley CJ. The cerebellum and Cognition: Evidence from Functional Imaging Studies. Cerebellum. In press 2011.
64. Swadlow HA, Waxman SG. Axonal conduction delays [online]. Dostupné z URL: http://www.scholarpedia.org/article/Axonal_conduction_delays.
Labels
Paediatric neurology Neurosurgery NeurologyArticle was published in
Czech and Slovak Neurology and Neurosurgery

2012 Issue 3
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
Most read in this issue
- Neurosyfilis
- Operační léčba syndromu tarzálního tunelu
- Oboustranná léze n. phrenicus manifestující se jako ortopnoe – kazuistiky tří případů
- Diagnostika a možnosti léčby Niemann-Pickovy choroby typ C