
Evoluce a evoluční teorie pro lékaře
V. Darwinovy teorie evoluce
Evolution and evolutionary theory for physicians: V. Darwin’s theories of evolution
“Darwin’s theory” is an inaccurate term. In reality there are five basic Darwinian theories of evolution. In addition to rejecting biblical literalism Darwin’s theories impeded and impede essentialism and finalism. Essentialism is a basis of transmutationism and transformationism, both antiquated theories. Darwin replaced essentialism by population thinking. So-called Modern synthesis (neo-Darwinism) evolved in the first half of the last century. Contemporary evolutionary genomics corroborates three of the six pillars of Modern synthesis:
- random heritable variations,
- uniformitarianism and
- the last universal common ancestor,
but refutes the next three pillars:
natural selection as the main driving force of evolution,
gradualism (evolution in „infinitesimally“ small steps), and
a single large Tree of life.
Key words:
Darwin’s five theories, six theoretical pillars of Modern synthesis (neodarwinism), contemporary evolutionary genomics.
Authors:
F. Koukolík
Authors‘ workplace:
Oddělení patologie a molekulární medicíny
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
Published in:
Prakt. Lék. 2010; 90(5): 268-273
Category:
Editorial
Overview
„Darwinova teorie“ je nepřesný pojem. Ve skutečnosti existuje pět základních teorií, jimiž Darwin evoluci vysvětloval. Kromě doslovné víry v Bibli přijetí Darwinovy teorie bránily a brání esencialismus a finalismus. Esencialismus je podkladem transmutacionismu a transformacionismu, překonaných teorií. Darwin nahradil esencialismus populačním myšlením. V průběhu první poloviny minulého století vznikla Moderní syntéza evoluční teorie – neodarwinismus. Současná evoluční genomika potvrdila tři ze šesti pilířů Moderní syntézy:
- náhodné dědičné variace jako materiální podklad evoluce,
- totožnost základních evolučních procesů v průběhu celé evoluce, a
- posledního univerzálního společného předka.
Zamítla však další tři:
- přírodní výběr jako dominantní hnací sílu evoluce,
- gradualismus, čili průběh evoluce v nepatrných krůčcích, a
- jednoduchou původní představu stromu života.
Klíčová slova:
Darwinových pět teorií, šest teoretických pilířů moderní syntézy (neodarwinismus), současná evoluční genomika.
Úvod
Pojem Darwinova teorie je stejně tradiční jako zjednodušující a nepřesný. Ve skutečnosti Darwin formuloval pět základních teorií (28):
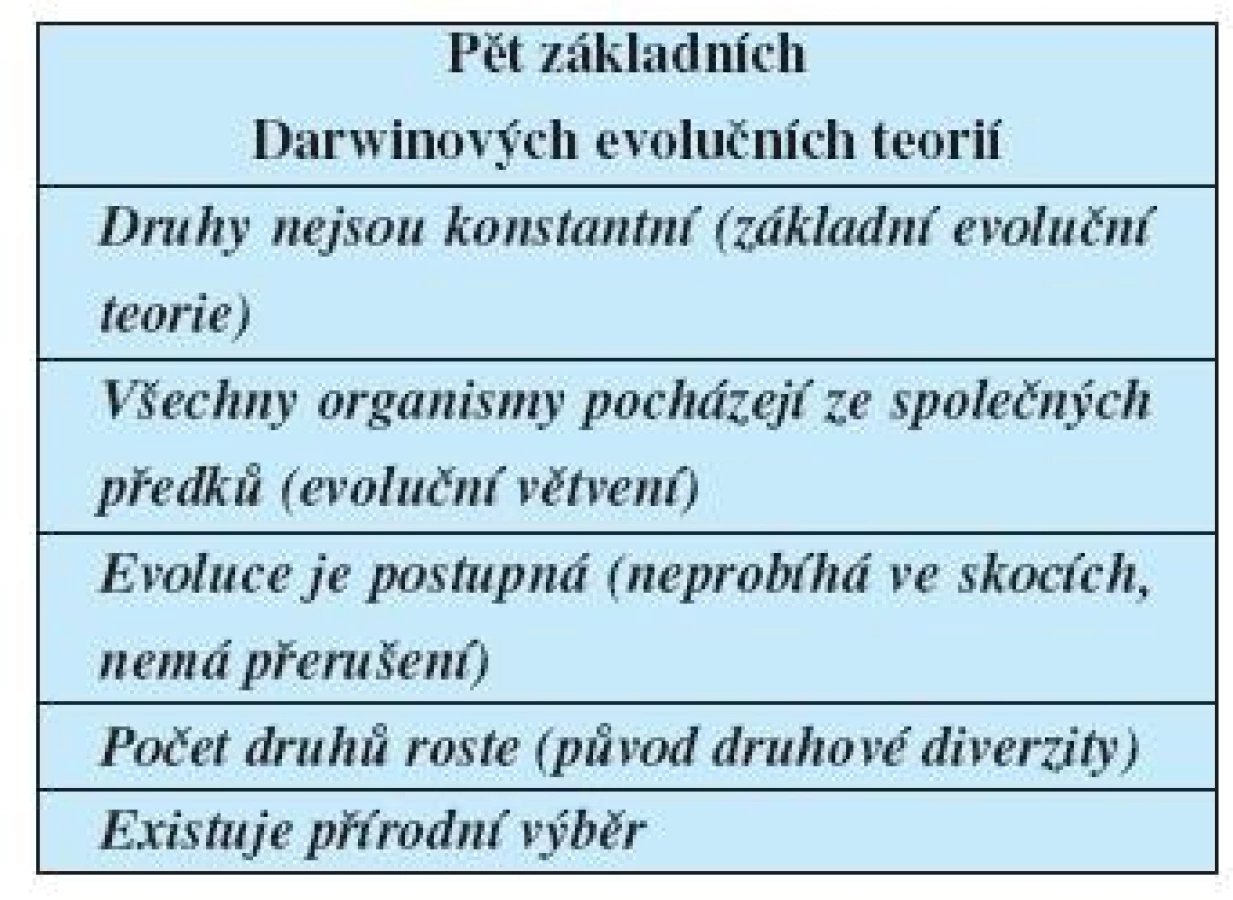
- druhy nejsou konstantní, což je základní evoluční teorie;
- všechny organismy pocházejí ze společných předků, to je teorie evolučního větvení;
- evoluce probíhá postupně, nikoli ve skocích nebo s diskontinuitami;
- počet druhů v čase a prostoru roste, to je důvodem jejich rozmanitosti;
- mechanismem evoluce je přírodní výběr.
Ranní evolucionisté se shodovali v názoru, že druhy nejsou konstantní. Ostatní čtyři teorie jak akceptovali, tak odmítali (tab. 1, 2).
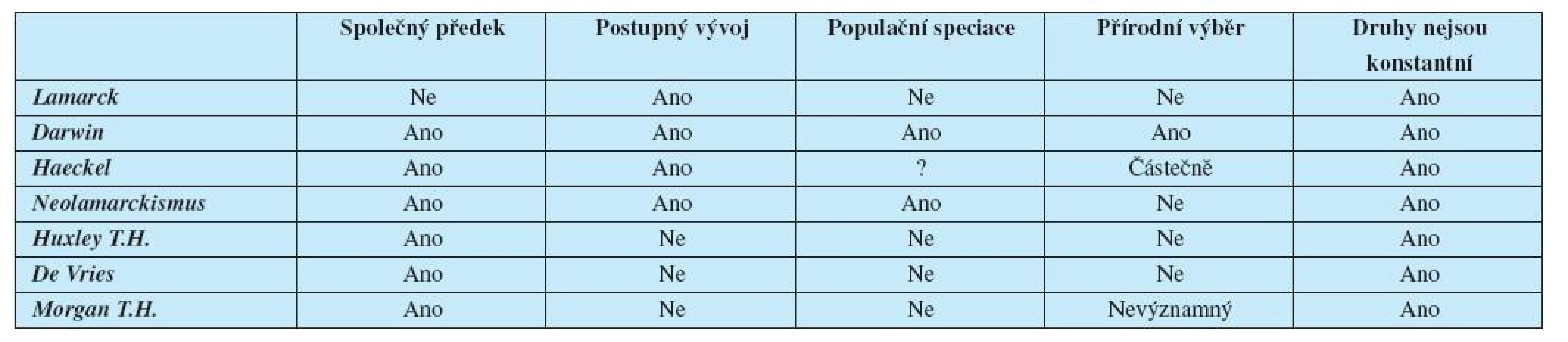
Přijetí Darwinovy teorie bránila a dodnes brání víra v doslovné znění Bible a filozofické představy, jejichž kořeny sahají do antiky – esencialismus a finalismus.
Esencialismus, neboli typologické myšlení, vytvořený pythagorejci a Platonem učí, že všechny přírodní jevy lze rozčlenit do tříd. Každá z nich je charakterizována svou „podstatou“ – esencí. Ta je konstantní a ostře vymezená vůči jiným esencím. V křesťanské představě byl každý druh života stvořen zvlášť, všechny žijící podoby druhu jsou potomci prvního páru stvořeného Bohem.
Esencialismus je podkladem zamítnutých teorií, které se pokoušely vysvětlit evoluci:
- transmutacionismu, a
- transformacionismu.
Ten má dvě podoby (tab. 3).
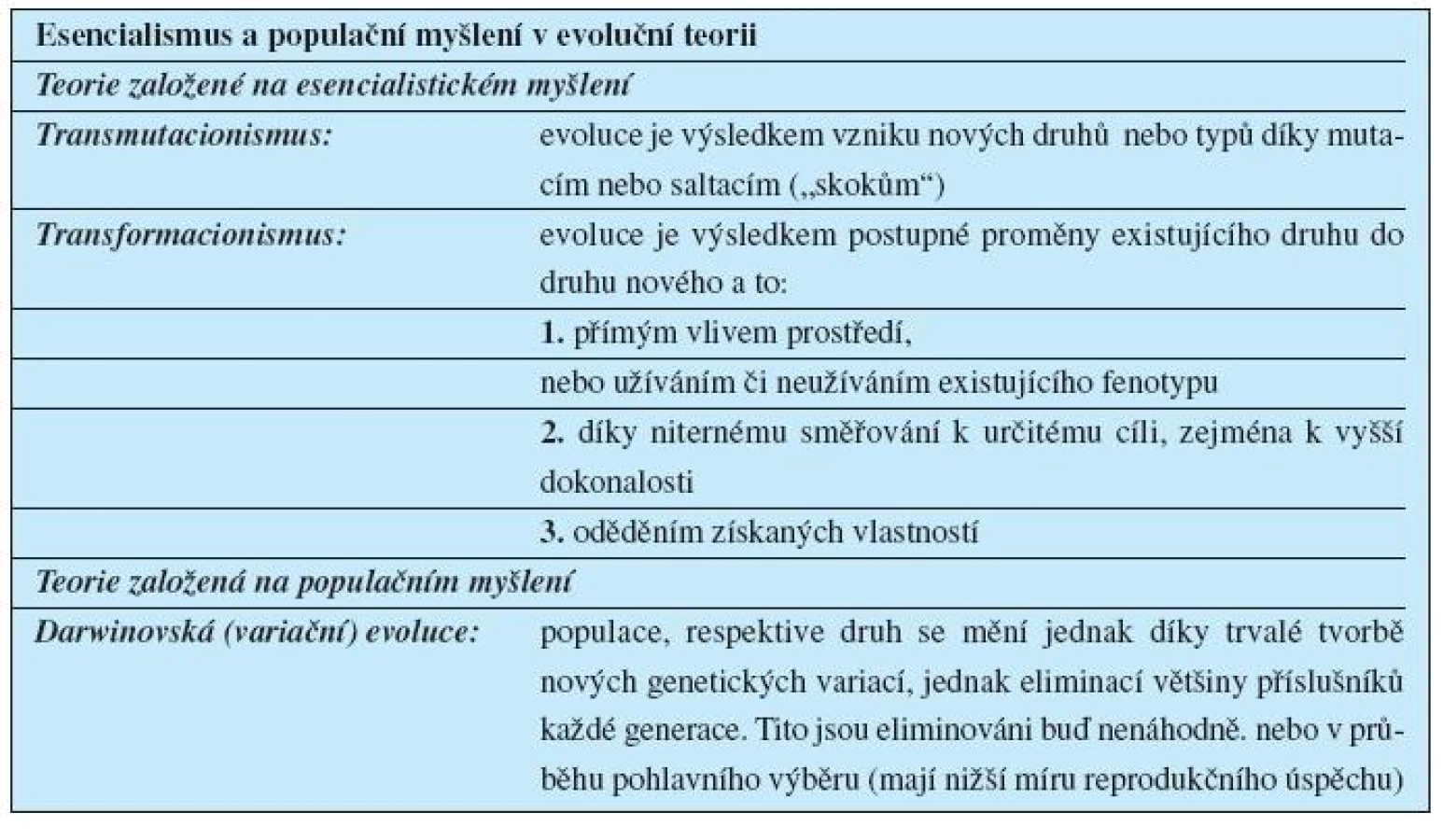
Transmutacionistický výklad působí intuitivně: současné i fosilní druhy dělají ostře oddělený dojem. Jejich mizení a objevování se v evoluci dělá dojem náhlé události. Skokovou, neboli saltacionistickou podobu transmutacionismu lze zamítnout jen tehdy, podaří-li se dobře vysvětlit „mezery“ v evolučních liniích.
Darwin namísto esencialismu zavedl populační myšlení. Živé organismy nejsou konstantní třídy, neboli typy, zato tvoří proměnlivé populace. Každý druh života je tvořen početnými místními populacemi. Každý jedinec v každé populaci se od jiného jedince v téže populaci odlišuje. Například lidský druh je v současnosti tvořen více než 6 miliardami vzájemně odlišných jedinců. Všichni jedinci v populaci nemohou mít stejný druh mutace současně. Nový druh tedy nemůže vzniknout náhle. Předpoklad, že by nový druh byl potomstvem „nadějné“ mutace (doslovně „nadějného monstra“), je málo pravděpodobný. Většina mutací je buď neutrálních, nebo poškozujících, či letálních.
Darwin neúnavně zdůrazňoval evoluci v malých krůčcích běžících dlouhou dobu.
Transformace díky vlivu prostředí předpokládá změnu genomu tlakem prostředí a přenos této změny do další generace – ať už jde o užívání, nebo neužívání. Známým příkladem pro užívání mělo být prodlužování krku u žiraf, příkladem zániku struktur neužíváním mělo být zanikání očí u jeskynních živočichů žijících ve tmě.
Experimentální genetika a molekulární biologie však dokázaly, že se geny přímým vlivem prostředí nemění.
Finalismus
je teorie, kterou lze stopovat k Aristotelovi. Živý svět podle něj směřuje k větší dokonalosti. Jde tedy o víru v konečný účel/smysl vesmíru a života. Evoluce má směřovat od nižšího k vyššímu, od jednoduchého ke složitému, od nedokonalého k dokonalému.
Transformace díky niternému „programu“ zdokonalování se nazývá ortogeneze. V průběhu evoluce nemají vznikat nové druhy, zato se zdokonalují ty staré. „Programy“ tohoto druhu nalezeny nebyly. Jestliže by existovaly, měla by být evoluce víceméně přímočará, což není. Evoluční linie mění směr. Může dojít i ke zvratu k primitivnějším podobám života, příkladem jsou oči a zbarvení živočichů žijících trvale v jeskyních, kde je tma.
Moderní syntéza evoluční teorie a genomika
Darwin neměl představu o mechanismu dědičnosti. Objasňování genetického základu evoluce začalo až po znovuobjevení Mendelových zákonů počátkem minulého století klasickými pracemi Fisherovými, Wrightovými a Haldaneovými. Během první poloviny minulého století vznikla zejména na základě prací Dobzhanského, Huxleových, Mayrových a Simpsonových Moderní syntéza evoluční biologie (neodarwinismus), která nabyla zralou podobu v r. 1959, v Chicagu, u příležitosti stého výročí Darwinova Vzniku druhů (35). Neodarwinismus by se dal nazvat darwinismem očima genetiky.
V průběhu následného půlstoletí proběhl další zlom – jeho název by mohl znít evoluční biologie ve světle genomiky (22). Šest teoretických pilířů Moderní syntézy změnila v následujících desetiletích a zejména v posledních letech genomika způsobem, který jeden z mých studentů přirovnal změně Newtonovy fyziky na Einsteinovu.
Základní kroky vývoje evoluční teorie v průběhu této doby jsou:
- hlubší pochopení molekulární evoluce a fylogeneze,
- vznik Kimurovy, Jukesovy a Kingovy neutrální teorie evoluce,
- teorie „sobeckého genu“, objev mobilních elementů a DNA „haraburdí“ (junk DNA),
- evoluce zdvojením genů a genomu,
- popis vedlejších výtvorů evoluce (tzv. spandrelů), exaptace, „fušování“ (tinkering) a kritika „panglosovského“ paradigmatu evoluce,
- hlubší pochopení evoluce mikrobů a virů,
- objev endosymbiózy.
Klasická fylogeneze se opírala o morfologii, porovnávala například kostry živočichů nebo architekturu květů. V šedesátých letech minulého století bylo zjištěno, že sekvence aminokyselin cytochromu c a globinů u živočichů, kteří jsou vzájemně vývojově značně vzdálení, jsou vysoce konzervované. Později byla doložena konzervace proteinů od bakterií k savcům a objevil se pojem molekulárních hodin: relativně konstantní evoluce sekvencí, aniž by to mělo funkční důsledky.
V roce 1987 Woese et al. (37) doložil konzervaci sekvencí RNA ve všech buněčných podobách života i jejich vhodnost pro fylogenetickou analýzu a objevil třetí doménu života – archea. Od této doby lze datovat vývoj podstatně hlubšího pochopení evoluce mikrobů a virů, o nichž sice evoluční teorie předpokládala, že jsou v kořenech „stromu života“, nicméně neměla nástroje, jimiž by se dala dobře zkoumat.
Neutrální teorie molekulární evoluce (17, 18)
má zato, že většina mutací, které se v průběhu evoluce udrží, je selekčně neutrálních. Jejich fixace je tedy výsledek náhodného driftu ( v této souvislosti obtížně převoditelný anglický pojem označující náhodné unášení proudem). Z teorie plyne, že se genové sekvence vyvíjejí způsobem popsaným hypotézou molekulárních hodin. „Nadějná monstra“ jsou natolik vzácná, že je není nutné brát v úvahu.
V dané souvislosti se mluví o očišťující selekci (purifying) která ponechává neutrální mutace, zato vyřazuje poškozující mutace. Očišťující charakter selekce je hlavní odlišností proti darwinovské selekci, ta je pozitivní.
Teorie neutrální selekce prošla dalším vývojem. V současnosti se nazývá téměř neutrální teorií. Mutace, které jsou téměř neutrální v době své fixace, jsou považovány za zdroj variability, která může být přírodním výběrem užita při změněných podmínkách, což je potenciálně významné pro makroevoluci, za kterou se obvykle považuje evoluce nad úrovní druhu.
Počínaje 60. léty minulého století se dále zjistilo, že organismy, jejichž fenotypická složitost je stejná, mají genomy, jejichž velikost se výrazně liší. Výsledkem byla teorie, podle níž z hlediska organismu může být většina jeho DNA nadbytečné „haraburdí“. V souvislosti s tímto objevem vznikla teorie sobeckého genu(2, 32, 4), podle níž podstatná část genomu vyšších organismů je dána replikací různých podob „sobeckých sekvencí“. Dawkinsova představa byla základním odchýlením od klasického přírodního výběru, jenž probíhá na úrovni jedince. Podle Dawkinse jsou cílem selekce geny.
Významným objevem datujícím se do 40. let minulého století jsou mobilní elementyjimž se dříve říkalo skákající geny. Prvně byly popsány u kukuřice, později u živočichů (1). Mobilní elementy často mění v genomu svou polohu. Genomy jsou tedy značně dynamické.
Ohno (31) má zato, že jádrem evoluce genů (a organismů) je zdvojování částí chromozomů. Zdvojení celého genomu se mělo objevit v rané evoluci strunatců. Zdvojování genů má být základní cestou evoluce nových funkcí. Na jednu ze dvou kopií genu totiž nemá vliv očišťující selekce.
Kritiky důsledně adaptacionistické představy evoluce, podle níž je „všechno“ výsledkem adaptací, se v proslulé práci z roku 1979 nazvané Spandrels of San Marco stali Gould a Lewontin (12).
Spandrel je označení zdiva mezi oblouky gotické katedrály. Spandrel je „vedlejší“, nicméně nutný „produkt“ její stavby. Stavitelé katedrál tyto „volné“ prostory následně zdobili. Gould a Lewontin (12) toto vševysvětlující pojetí evoluce odmítají. Označují je jako „panglosovské“. (Doktor Panglos je postava Voltairova románu Candide. Voltaire se doktorem Panglosem posmíval německému filozofovi G.W. von Leibnizovi (1646 –1716), jenž viděl náš svět jako nejlepší z možných světů, v němž vše, co se děje, a to včetně nejhorších obecných a osobních katastrof, se děje k jeho vylepšení). Oba autoři oprávněně poukazují na nutnost vzít při analýze evoluce v úvahu kromě adaptací náhodný drift a další faktory. Využití „spandrelů“ nazval Gould pojmem exaptace a považuje ho za významný evoluční mechanismus (13).
Endosymbióza,
vznik některých organel eukaryotických buněk z bakterií byl nápad z konce 19. století. Dlouhou dobu šlo o čistou spekulaci. Teprve v šedesátých letech minulého století byl na základě genomu chloroplastů a mitochondrií podán důkaz (34, 27). Mitochondrie pocházejí z alfa-proteobakterií (14).
Současný vývoj
Současný vývoj evoluční teorie je dán znalostí celých genomů: několika tisíc virových, téměř tisícovky genomů bakterií a archeálních a přibližně stovky genomů eukaryot (7, 24). Porovnávání celých genomů umožňuje rozlišovat ortologní geny pocházející z jednoho společného předka od paralogních genů vzniklých zdvojením. Intenzivně se rozvíjí metagenomika, studující genetický materiál získaný z nějakého prostředí, například z úst, střeva, mořské nebo odpadní vody, nikoli jen z laboratorních kultur.
Metagenomika, byť se teprve rodí, ozřejmila neuvěřitelnou rozmanitost podob života ve všech studovaných prostředích. V mořských habitatech jsou nejpočetnějšími představiteli života viry. Viry jsou rovněž geneticky nejrozmanitější podoba života. Pilotní metagenomický projekt vedený Craigem Venterem našel ve vodě Sargasového moře DNA téměř 2 000 různých druhů života včetně 148 do té doby neznámých bakterií (34). Lze říci že vzniká nový, syntetický obor – evoluční systémová biologie(20).
Pravděpodobně nejzajímavější objev současného vývoje je paradoxně působící evoluční stabilita, neboli konzervování genových sekvencí na jedné straně, a proměnlivost složení genů a architektury genomu na straně druhé. Například sekvencování genů mořské sasanky a vločkovce (trichoplax), asi třímilimetrového, průsvitného primitivního tvora tvořeného jen dvěma buněčnými vrstvami, dorzální a ventrální, dokázalo při porovnání s ptáky a savci rozsáhlou genovou konzervaci. Živočišné geny jsou tedy staré stovky milionů let. Porovnání genomů rostlin, hub a prokaryot došlo ke stejnému závěru (21). Předpokládá se, že nositelem předchůdců stovek současných genů byl poslední univerzální společný předek (LUCA – last universal common ancestor; 29).
Koonin (22) má za to, že sekvence mnoha genů kódujících transkripci, translaci, replikaci a základní metabolické dráhy, jsou předmětem očišťující selekce po celou dobu vývoje života – asi 3,5 miliardy let. Podobně konzervativně se chovají introny – prokazuje se 25–30 % konzervace ortologních genů od rostlin po strunatce (9). Vysoký podíl polohy intronů sdílí například mořská sasanka a vločkovec se savci. Na druhé straně platí, že „téměř univerzální geny všeho života“ jejichž počet se odhaduje kolem 70, tvoří jen zlomek celkového počtu genů, a to kolem 10 % v nejmenších známých genomech, obvykle však kolem 1 %.
Proti tomuto stabilnímu genomickému jádru života stojí vysoce dynamický obraz genomické evoluce ze ztrátami řady genů horizontálním přenosem a zdvojováním jiných genů. S výjimkou operonů, to jsou skupiny genů pod kontrolou jednoho regulačního signálu, respektive promotoru, je organizace genů u prokaryot dána rozsáhlými náhodnými přesuny. Porovnání genomické organizace archea a bakterií prokázalo zisk podstatné části genomu některých prokaryot horizontálním přenosem (10), nějakým druhem vzájemné směny. Jestliže bakterie a archea sdílejí habitat, dochází mezi nimi také k horizontálnímu přenosu genů. U mnohobuněčných eukaryot jsou somatické buňky odděleny od zárodečných.
Horizontální přenos genů je však prokázán mezi endosymbiotickými bakteriemi a genomem jejich hostitele. Naproti tomu existuje častá směna genů mezi bakteriemi a jednobuněčnými eukaryoty a mezi jednobuněčnými eukaryoty vzájemně.
Je také nutné mít na mysli endosymbiózu popsanou v předešlém sdělení: eukaryota jsou genetické chiméry, které nesou geny bakterií a archea (8). Ze současného poznání plyne základní zobecnění: genomy všech podob života jsou soubory genů, které mají vzájemně odlišnou evoluční historii (22, 23). Z toho plyne změna základní evoluční představy, stromu života (ToL – Tree of Life), protože topologie jednoho evolučního stromu, ani odpovídající topologie většího počtu stromů malého počtu vysoce konzervovaných genů nereprezentuje vývojovou historii většiny genů (5).
V současnosti jsou navrhovány dvě obměněné podoby stromu života:
1. strom,
tvořený vysoce konzervovanými geny tvořící ústřední směr evoluce. Horizontální přenos genů včetně endosymbiózy vytváří horizontální propojení jednotlivých větví stromu.
2. komplexní síť,
v níž existují jednak fáze charakterizovatelné evolucí podobné větvení stromu včetně horizontálního propojení větví, jednak fáze „velkých třesků“, rychlých horizontálních výměn genetické informace (22, 23). Sekvence odvozené od mobilních elementů, zvláště retrotransposony, tvoří kolem 40 % savčích genomů (11).
Pro druhou možnost svědčí prohlubují se znalost „mobilomu“ (též virosféry, nebo světa virů), což je označuje vzájemně propojená genetická zásobárna tvořená viry a dalšími „sobeckými“ replikony, to jsou genetické elementy, které nekódují celistvý translační systém, například různé druhy plazmidů a transposonů (19).
Srovnávací genomika a metagenomika našly rozsáhlý, dynamický vzájemně provázaný svět „sobeckých“ replikonů v interakci s genomy buněčných podob života. Replikony přispěly zásadním způsobem v průběhu evoluce ke tvorbě chromozomů. Bakteriální chromozomy lze obrazně považovat za „ostrovy stability“ v moři mobilních elementů. Intragenomická pohyblivost „sobeckých“ transposonů v genomu eukaryot je rozsáhlá. Je pravděpodobné, že evoluční přechody souvisí s „výbušnými“, to znamená rychlými a rozsáhlými propagacemi mobilních elementů (26).
Zásadní ochranu proti virům a některým mobilním prvkům představuje RNA interference (RNAi). Ta degraduje virovou RNA. K manipulaci genové exprese však může tento mechanismus užívat jak hostitel, tak virus. Hostitel může kódovat mikroRNA, pro kterou je virová sekvence cílem. Viry však vyvinuly početné adaptace, které mohou RNAi potlačit, nebo se jí vyhnout. RNAi může chránit buňky proti mobilním elementům, a to jak degradací jejich transkriptů, tak blokováním jejich exprese formováním heterochromatinu. Geny kódující protivirovou, stejně jako RNAi likvidující mobilní elementy, se vyvíjejí rychle – stejně rychle se však vyvíjejí virové supresory RNA. Mezi hostiteli a parazity tedy probíhají evoluční závody ve zbrojení. V dlouhých evolučních obdobích vývoje mnohobuněčných podob života se RNAi opakovaně zdvojovaly i ztrácely (30).
Darwinův předpoklad původu všech současných podob života z jediného společného předka srovnávací genomika dokládá. Stovky konzervovaných genů ukazují, že by tímto společným předkem mohl být LUCA, last common universal ancestor, poslední společný univerzální předek. Rekonstrukce ukazují, že LUCA měl celistvý translační systém, který byl blízký jednouché podobě současné translace, takže sestával z přibližně stovky RNA a proteinových molekul, základního systému transkripce a základních metabolických drah, například biosyntézy purinových a pyrimidinových nukleotidů.
Na druhé straně se mechanismus replikace DNA u bakterií, archea a eukaryotů vzájemně odlišuje, podobně jako se odlišují jejich membránové lipidy a enzymy odpovídající za tvorbu membrán, což svědčí pro to, že kódující geny nejsou ortologické. Existuje tedy možnost, že LUCA nebyl buňka, ale spíše zásobárna (pool) genetických elementů, které se replikovaly a exprimovaly odlišně a moderní replikace DNA a mebránové systémy se za předpokladu symbiogenetického vzniku eukaryot vyvinuly nezávisle dvakrát. Koonin (22) proto zavádí pojem LUCAS, last universal common ancestral state. Odlišná představa říká, že LUCA byl buňkou moderního typu, která mohla být vybavena bakteriální nebo archeální podobou replikace DNA, případně jejich směsí.
Viry a prokaryota mají „natěsnané“ genomy tvořené geny kódujícími strukturální RNA a proteiny, nekódující oblasti tvoří až na výjimky 10–15 % genomické DNA. Podobně vypadají genomy jednobuněčných eukaryot. Až na pozoruhodné výjimky, mezi než patří například proslulá japonská ryba fugu, je situace u mnohobuněčných eukaryot, včetně lidí, opačná: většinu genomické DNA tvoří nekódující úseky. Jev lze vysvětlit očišťující selekcí, která ovlivňuje populaci úměrně k její efektivní velikosti (to je podíl populace schopný mít potomstvo). Otázka, do jaké míry je genetické „haraburdí“ (junk DNA), tedy nekódující součásti genomu, skutečně haraburdím, je však stále otevřená (22, 23).
Většina genomů buněčných podob života s výjimkou malých genomů obligatorních parazitů obsahuje paralogní geny svědčící pro předchozí zdvojení. Ohnovo (31) zdvojování genů se považuje za hlavní mechanismus evoluce, protože touto cestou vznikají nové geny (22). Předpokládá se, že si vývoj zdvojených genů podrží část funkcí původního genu, zbytek různým tempem zaniká (25).
Zdvojování genů se objevuje ve všech vývojových liniích, jeho tempo však není jednotné. Předpokládá se, že diferencované zdvojování skupin Hox (homeobox) genů a dalších regulátorů vývoje bylo klíčově významné pro vývoj živočišných kmenů (15). Za skokové události se považují zdvojení celého genomu, po němž následuje rozsáhlá ztráta genů prokázaná jak u kvasinek, tak u rostlin a strunatců. Předpokládá se, že tato událost měla základní význam pro primární radiaci strunatců (3).
Současné metody umožňují rekonstruovat evoluční historii proteinových komplexů včetně určování jejich stáří. Proteinové komplexy vznikaly již v časné evoluci a vyvíjely se jako koherentní jednotky. Z modelu plyne v jaké míře se jejich vzniku účastnilo zdvojování genů: týká se přibližně čtvrtiny orthologních komplexů (38).
Průkaz evolučního významu genového a celogenomového zdvojování zamítá klasickou Darwinovu gradualistickou představu. Zdvojení genomu lze považovat za událost skokovou (saltatorní). Zdvojení genů a následná často rychlá diverzifikace paralogních genů jako způsobu vzniku nových genů vede k metafoře evoluce coby „fušéra“ (tinkerer), jenž tvoří nové nástroje tím, že okopíruje staré a kopie následně upraví.
Komplexita
Moderní syntéza (neodarwinismus) předpokládala, že komplexita organismů – mnohobuněčné organismy jsou složitější než jednobuněčné – je výsledkem početných adaptivních proměn daných přírodním výběrem. Měl tomu odpovídat počet nukleotidů nesoucích funkčně relevantní informaci. Genomy mnohobuněčných organismů jsou složitější než genomy jednobuněčných organismů. Klíčovým mechanismem komplexity složitějších organismů měl být alternativní splicing,jehož prostřednictvím může vznikat mnohonásobně vyšší počet proteinů, než je odpovídajících genů. Jestliže má Homo sapiens sapiens přibližně 20 000 genů a bakterie 10 000 genů, neplyne z toho, že je lidský proteom dvakrát komplexnější než bakteriální – právě díky alternativnímu splicingu (33).
Dokazuje se však, že alternativní splicing se jako adaptace nevyvinul. Rekonstrukce prokazují, že hustota intronů vývojově starých eukaryot byla srovnatelná s jejich hustotou v genomech bohatých na introny, jako je tomu u obratlovců. Početné „omyly“ vznikající při splicingu odstraňuje NMD systém (nonsense-mediated decay; 16), konzervovaný u všech eukaryot. Alternativní splicing se objevil pravděpodobně jako genomická „porucha“.
Ne-adaptacionistická populačně genetická teorie evoluce genomické komplexity (26)
předpokládá, že genetické změny vedoucí k růstu komplexity, například zdvojování genů nebo inzerce intronů jsou mírně poškozující. Jejich výskyt se v populaci udržuje jen při slabé očišťující selekci. Jestliže je síla očišťující selekce úměrná velikosti efektivní populace, poroste genomická komplexita jen v době průchodu populace „úzkým hrdlem“, tedy jakmile se podíl efektivní populace dostane mezi přežití druhu. Genomická komplexita v tomto případě není adaptací, ale je výsledkem evolučně neutrálních procesů v případě neefektivní očišťující selekce. Uvedený princip však má četné výjimky (22).
Výsledek genomické evoluce závisí na rovnováze mezi tlakem očišťující selekce, jejíž mohutnost je závislá
- na velikosti populace a četnosti mutací,
- na rekombinačních procesech,
- na aktivitě „sobeckých“ elementů, a
- na adaptaci na specifické habitaty.
Evoluce genomické komplexity závisí na kombinaci neutrálních (náhodných, stochastických) a adaptivních procesů (23). Kromě toho se prokazuje, že v průběhu 3,5 miliardy let života setrvalý vývoj komplexity od nižší k vyšší, a to jak komplexity organizační, tak genomické, neexistoval. O „pokroku“ coby součásti evoluce mluvit nelze (22).
MIM hypotéza
(Mistranslation-induced Misfolding)
říká, že tempo vývoje nových sekvencí je určováno tempem translačních událostí. Jinak řečeno, jde o selekci odolnosti vůči chybnému prostorovému uspořádání proteinů (protein misfolding; 6). Tyto proteiny jsou toxické a vedou k zániku buněk, jak v praxi ukazují četná neurodegenerativní onemocnění včetně prionových
Souhrn: moderní syntéza a evoluční genomika
Co říká o šesti pilířích moderní syntézy (MS) evoluční genomika (EG) (tab. 4)?
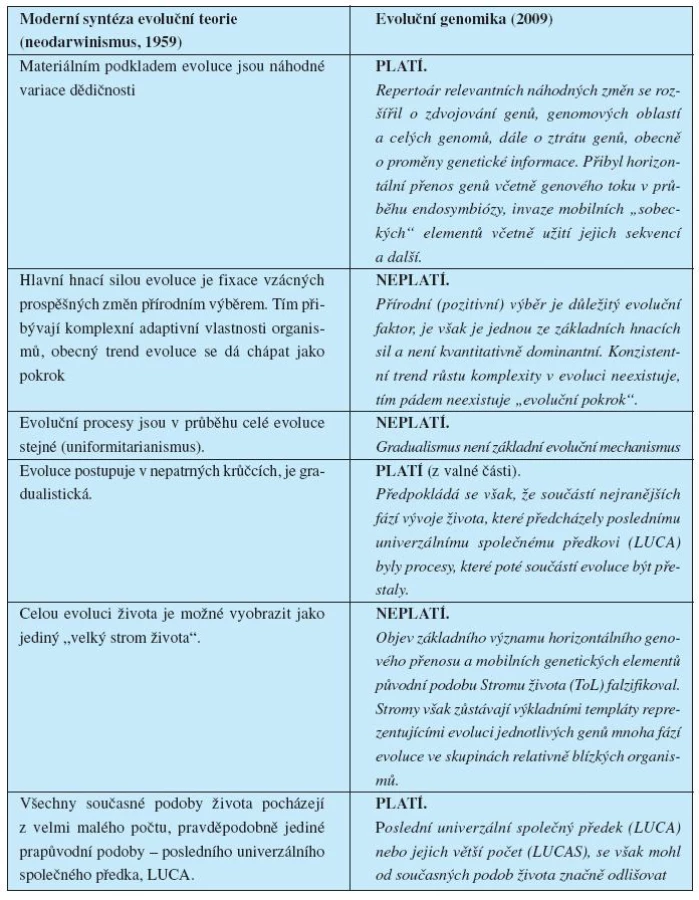
1. pilíř
MS: materiálním podkladem evoluce jsou náhodné variace dědičnosti
EG: Stojí. Repertoár relevantních náhodných změn se rozšířil o zdvojování genů, genomových oblastí a celých genomů, dále o ztrátu genů, obecně o proměny genetické informace. Přibyl horizontální přenos genů včetně genového toku v průběhu endosymbiózy, invaze mobilních „sobeckých“ elementů včetně užití jejich sekvencí a další.
2. pilíř
MS: Hlavní hnací silou evoluce je fixace vzácných prospěšných změn přírodním výběrem. Tím přibývají komplexní adaptivní vlastnosti organismů, obecný trend evoluce se dá chápat jako pokrok.
EG: Spadl. Přírodní (pozitivní) výběr je důležitý evoluční faktor, je však jen jednou ze základních hnacích sil a není kvantitativně dominantní. Evoluce je výsledkem neutrálních (stochastických) procesů kombinovaných s očišťující selekcí, která odstraňuje „nepovedené“ výtvory. Genomická komplexita je výsledkem slabé očišťující selekce v malých populacích, výsledkem adaptací není. Konzistentní trend růstu komplexity v evoluci neexistuje, tím pádem neexistuje „evoluční pokrok“.
3. pilíř
MS: Variace fixované přírodním výběrem jsou „infinitezimálně malé“. Evoluce postupuje v nepatrných krůčcích, je gradualistická.
EG: Spadl. Zdvojení genu, delece, akvizice delších oblastí, přeskupení, natož zdvojení celého genomu nebo endosymbióza nejsou infinitezimálně malá variace. Gradualismus není základní evoluční mechanismus.
4. pilíř
MS: „Uniformitarianismus“ – evoluční procesy jsou v průběhu celé evoluce stejné.
EG: Stojí, z valné části. Předpokládá se však, že součástí nejranějších fází vývoje života, které předcházely poslednímu univerzálnímu společnému předkovi (LUCA), byly procesy, které poté součástí evoluce být přestaly. Velké evoluční přechody, jejichž příkladem je vznik eukaryot, lze vyložit unikátními událostmi jako je endosymbióza.
5. pilíř
MS: Celou evoluci života je možné vyobrazit jako jediný „velký strom“.
EG: Spadl. Objev základního významu horizontálního genového přenosu a mobilních genetických elementů původní podobu Stromu života (ToL) falzifikoval. Stromy však zůstávají výkladními templáty reprezentujícími evoluci jednotlivých genů mnoha fází evoluce, ve skupinách relativně blízkých organismů. Možnost záchrany Stromu života coby centrálního vývojového směru evoluce zůstává.
6. pilíř
MS: Všechny současné podoby života pocházejí z velmi malého počtu, pravděpodobně jediné prapůvodní podoby – posledního univerzálního společného předka, LUCA.
EG: Stojí. Srovnávací genomika dokládá společného předka buněčných podob života. Poslední univerzální společní předek (LUCA) nebo jejich větší počet (LUCAS), se však mohl od současných podob života značně odlišovat (22).
MUDr. František Koukolík, DrSc.
Oddělení
patologie
a molekulární
medicíny
Národní
referenční laboratoř
prionových
chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská
800
140
59 Praha 4 Krč
E-mail:
frantisek.koukolik@ftn.cz
Sources
1. Clintock, B. The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. USA 1950, 36, p. 344-355.
2. Dawkins, R. The selfish gene. Oxford: Oxford University Press, 1976.
3. Dehal, P, Boore, J.L. Two rounds of whole genome duplication in the ancestral genome. PLoS Biol. 2005, 3: e314.
4. Doolitle, W.F., Sapienza, C. Selfish genes , the phenotype paradigm and genome evolution. Nature 1980, 284, p. 601-603.
5. Doolitle, W.F., Baptiste, E. Pattern pluralism and the Tree of Life hypothesis. Proc. Natl. Acad. Sci. USA 2007, 104, p. 2043-2049.
6. Drummond, D.A., Bloom, J.D., Adami, C. et al. Why highly expressed proteins evolved slowly. Proc. Natl. Acad. Sci. USA 2005, 102, p. 14338-14343.
7. Eisen, J.A., Fraser CM. Phylogenomics: intersection of evolution and genomics. Science 2003, 300, p. 1706-1707.
8. Embley, T.M., Martin, W. Eukaryotic evolution, changes and challenges. Nature 2006, 440, p. 623-630.
9. Fedorov, A., Merican, A.F., Gilbert, W. Large-scale comparison of intron positions among animal plant and fungl genes. Proc. Natl. Acad. Sci. USA 2002, 99, p.16128-16133.
10. Gogarten, J.P., Townsend, J.P. Horizontal gene transfer, genome inovation and evolution. Nat. Rev. Microbiol. 2005, 3, p. 679-687.
11. Goodier, J.L., Kazazian, H.H. Jr. Retrotransposons revisited. The restraint and rehabilitation of parasites. Cell 2008, 135, p. 23-35.
12. Gould, S.J., Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond. B. Biol. Sci. 1979, 205, p. 581-598.
13. Gould, S.J. The exaptive excellence of spandrels as a term and prototype. Proc. Natl. Acad. Sci. USA 1997, 94, p.10750-10755
14. Gray, M.W., Burger, C., Lang, B.F. The origin and early evolution of mitochondria. Genome Biol. 2001, 2(6), reviews1018.1–1018.5.
15. Hoegg, S. Meyer, A. Hox clusters as models for vertebrate genome evolution. Trends Genet. 2005, 201, p. 421-424.
16. Jaillon, O., Bouhouche, K., Gout J,F. et al. Translational control of intron splicing in eucaryotes. Nature 2008, 451, p. 359-362.
17. Kimura, M. Evolutionary rate at the molecular level. Nature 1968, 217, p. 624-626.
18. King, J.L., Jukes, T.H. Non-darwinian evolution. Science 1969, 164, p. 788-798.
19. Koonin, E.V., Senkevich, T.G., Dolja, V.V. The ancient virus world and the evolution of cells. Biol. Direct. 2006, 1, p. 29.
20. Koonin, E.V., Wolf, Y.I. In: Pagel M., Pomiankowski A (eds.). Evolutionary genomic and proteomics. Sunderland MA: Sinauer Ass. Inc., 2008 A, p. 11-25.
21. Koonin, E.V., Wolf, Y.I. Genomics of bacteria and archaea: the emerging generalizations after 13 years. Nucleic. Acid. Res. 2008, 36, p. 6688-6719.
22. Koonin, E.V. Darwinian evolution in the light of genomics. Nucleic. Acid. Research 2009, 37, p. 1011-1034.
23. Koonin, E.V. Evolution of genome architecture. Int. J. Biochem. Cell Biol. 2009, 41, p. 298-306.
24. Liolios, K., Mavromatis, K., Tavernarakis, N. et al. The Genomes On Line Database (GOLD) in 2007: status of genomic and metagenomic projects and their associated metadata. Nucleic. Acid. Res. 2008, 36, D475-479.
25. Lynch, M., Katju, V. The altered evolutionary trajectories of gene duplicates. Trends Genet. 2004, 20, p. 544-549.
26. Lynch, M. The origin of genome architecture. Sunderland MA: Sinauer Associates, 2007.
27. Martin, W., Hoffmeister, M., Rotte, C. et al. An overview of endosymbiotic models for the origin of eukaryotes, their ATP-producing organelles (mitochondria and hydrogenosomes) and their heterotrophic lifestyle. Biol. Chem. 2001, 382, p. 1521-1539.
28. Mayr, E. What evolution is. New York: Basic Books, 2001.
29. Mushegian, A. Gene content of LUCA, the last universal common ancestor. Front. Biosci. 2008, 13, p. 4657-4666.
30. Obbard, D.J., Gordon, K.H.J., Buck, A.H. et al. The evolution of RNAi as a defense against viruses and transposable elements. Phil. Trans. R. Soc. B 2008, 364(1513), p. 99-115.
31. Ohno, S. Evolution by gene duplication. Berlin-Heidelberg-New York: Springer-Verlag, 1970.
32. Orgel, L.E., Crick, F.H. Selfish DNA: the ultimate parasite. Nature 1980, 284, p. 604-607.
33. Park, J.W., Graveley, B.R. Complex alternative splicing Exp. Med. Biol. 2007, 623, p. 50-63.
34. Sagan, L. The origin of mitosing cells. J. Theor. Biol. 1967, 14, p. 255-274.
35. Tax, S., Callender, C. (Eds.) Evolution after Darwin; the University of Chicago Centennial. Chicago: University of Chicago Press, 1960.
36. Venter, J.C., Remington, K., Heidelberg, J.F. et al. Environmental genome shotgun sequencing of the Sargasso sea. Science 2004, 304, p. 66-74.
37. Woese, C.R. Bacterial evolution. Microbiol. Rev. 1987, 51, p. 221-271.
38. Yosef, N., Kupiec, M., Ruppin, E. et al. A complex-centric view of protein network evolution. Nucleic. Acid. Res. 2009, 37(12), e88.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2010 Issue 5
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
Most read in this issue
- Alimentární infekce a intoxikace bakteriálními toxiny
- Existuje bisexualita?
- Účinná hemostáza při bezkrevních chirurgických operacích
- Posuzování zdravotního stavu žadatelů pro účely náhradní rodinné péče