
Neurobiologie dospívání
Neurobiology of adolescence
Adolescence is a transition phase between childhood and adulthood, a time of learning and adjustment, particularly in the setting of long-term goals. It also is a time when youths are discovering how to navigate new social challenges and are adjusting to a large number of physical, cognitive and emotional changes within themselves. Throughout adolescence continues dramatic structural and functional brain development. Developmental neuroimaging studies are inconsistent with a simple model of frontal cortical immaturity. Changes in social and affective processing during adolescence – changes in so called „social brain“ – may confer greater flexibility in adjusting one´s motivations and goals in rapidly changing contemporary social world.
Keywords:
adolescence – frontal cortical immaturity – social brain
Authors:
F. Koukolík
Authors‘ workplace:
Primář: doc. MUDr. Radoslav Matěj, PhD.
; Národní referenční laboratoř prionových chorob
; Thomayerova nemocnice, Praha
; Oddělení patologie a molekulární medicíny
Published in:
Prakt. Lék. 2013; 93(6): 247-253
Category:
Editorial
Overview
Adolescence je přechodné období mezi dětstvím a dospělostí, doba učení a přizpůsobování zejména při tvorbě dlouhodobých cílů. V této době mladí lidé objevují jak hledat správný směr v nových sociálních výzvách. Přizpůsobují se velkému počtu tělesných, poznávacích a emočních změn v sobě samých. Vývojové studie zobrazovacími metodami jsou v rozporu s jednoduchým modelem nezralosti čelních laloků. Změny zpracovávání sociálních a afektivních informací v průběhu dospívání – změny tzv. „sociálního mozku“ – mohou poskytovat vyšší míru pružnosti přizpůsobování osobních motivací a cílů v současném rychle se měnícím sociálním světě.
Klíčová slova:
dospívání – nezralost čelní kůry – sociální mozek
ÚVOD
Dospívání je přechodná fáze mezi dětstvím a dospělostí (13), jiná definice říká, že jde o periodu mezi začátkem puberty a dosažením stabilní a nezávislé role ve společnosti (48). Začíná mezi 9–12 rokem po narození, u dívek obvykle o 1–2 roky dříve než u chlapců.
Počátek puberty je dán kaskádou hormonálních změn (44). Kaskáda počíná hypotalamickou sekrecí hormonu uvolňujícího gonadotropiny (GnRH), uvolňuje se v pulzech během spánku. Tím se znovu nabudí osa hypotalamus – hypofýza – nadledvina, která byla prvotně aktivní v průběhu prenatálního a raně neonatálního období, v dětství je její činnost utlumená. Pulzy GnRH podnítí hypofýzu k sekreci FSH, hormonu stimulujícího folikuly a luteinizačního hormonu (LH). Ty stimulují ovaria a testes, které začnou secernovat pohlavní hormony estrogen, resp. testosteron.
Druhou neuroendokrinní osou dospívání je hypofyzární sekrece růstového hormonu, rovněž souvisí se spánkem.
Třetí, zatím nejméně prozkoumanou osou je uvolňování dehydroepiandrosteronu (DHEA) z nadledvin.
Hlavní hormony regulující tělesné změny a vznik sekundárních pohlavních znaků jsou testosteron, estradiol a DHEA. Pohlavní dimorfismus lze přičíst jak rozdílům v hladinách pohlavních hormonů, vyšší hladině estradiolu u dívek, testosteronu u chlapců, tak rozdílům distribuce a typu hormonálních receptorů v cílových tkáních. Mechanismus probouzející sekreci GnRH v pubertě není zcela jasný. Uplatňuje se leptin, hormon tvořený tukovými buňkami, jenž se podílí se regulaci příjmu a výdeje energie a na chuti k jídlu a kisspeptiny, neuropeptidy podmiňující vliv leptinu na GnRH neurony v hypotalamu.
Neurobehaviorální změny pozorované v průběhu lidské adolescence, například vyšší tendence ke zkoumání prostředí a změny ve zpracovávání odměny, jsou známy u různých zvířecích druhů (44, 45). Podobají se změnám v adolescentním lidském mozku. Schlegel a Barry (43), antropologové, dokládají existenci adolescence coby přechodného období mezi dětstvím a dospělostí studiem 186 lidských společností, včetně společností lovců – sběračů. V současné postindustriální společnosti se objevují první známky puberty mezi 9.–11. rokem, menarché je v USA ve 12 letech, takže puberta počíná dříve než v minulosti, naproti tomu dospělé role zaujímají dnešní mladí lidé často v polovině třetího decenia, tedy později než v minulých dobách. Adolescence se oproti minulosti prodloužila. Současní adolescenti prožívají delší období ve škole spolu se svými věkovými vrstevníky.
ZOBRAZOVACÍ METODY
Vyšetřování vývoje mozku v adolescenci rozsáhle užívá strukturální i funkční metody, zejména magnetickou rezonanci. Věkové rozmezí vyšetřovaných skupin bývá 8–25 let. Velikost a vývojová trajektorie rozdílů záleží na způsobu vyšetřování a na tom, zda a jak byly uvedené struktury vztaženy k rozdílům v celkovém objemu mozku a k věku. V průběhu dětství a dospívání se zvětšuje objem korové bílé hmoty. Korová šedá hmota, která odpovídá hustotě neuronů a počtu synapsí jakož i míře myelinizace intrakortikálních axonů, sleduje vývoj, který se popisuje křivkou ve tvaru písmene U. V různých mozkových oblastech probíhá vývoj různě a nadto různě u dívek a chlapců – dívky „předbíhají“ chlapce přibližně o 3 roky. Redukce korové šedé hmoty probíhá pomalu zejména v prefrontální dorzolaterální kůře a kůře na hranicích spánkového a temenního laloku (temporoparietální funkce, TPJ), změny pokračují ještě v prvních letech třetího decenia (18–21, 34).
Blakemoreová (5, 6) uvádí relativní růst objemu bílé hmoty a relativní ubývání šedé hmoty čelních a temenních laloků dospívajícího mozku jakož i růst frakční anisotropie bílé hmoty v metodě DTI (diffuse tensor imaging). Frakční anisotropie udává rozsah, v němž není difuze molekul vody v bílé hmotě ve všech směrech stejná, což se považuje za míru organizace neuronálních drah. Změny tohoto druhu jsou v průběhu adolescence nelineární: rychleji probíhají v rané adolescenci, pak zpomalují, v rané dospělosti se vyrovnávají.
Longitudinální studie ve věku 4–22 let prováděná ve dvouletých odstupech prokázala růst objemu šedé hmoty čelních laloků v pozdním dětství a rané adolescenci vrcholí ve věku přibližně 12 let. Následně její objem klesal. Podobný vývoj probíhá v kůře laloků temenních (18). Další longitudinální studie sledovala rovněž v dvouletých odstupech vývoj mozku v rozmezí 4–21 let věku. Měřeno denzitou šedé hmoty vyzrávaly nejprve senzorické a motorické korové oblasti, následovaly další korové úseky, a to ve směru od zadních k předním částem mozku. Jako poslední vyzrávala horní spánková kůra (22, 42). Mezi dětstvím a adolescencí se ztrácí šedá hmota v dorzální prefrontální kůře a v kůře temenní, ve stejných oblastech, zejména prefrontální přibývá hmota bílá. Ztráta denzity šedé hmoty jde na vrub intrakortikální myelinizace a růstu axonálního průměru, jinou interpretací je synaptická reorganizace (39). Růst objemu šedé hmoty v některých oblastech mozku v průběhu puberty, stejně jako pokles její denzity je tedy možné teoreticky přičítat kombinaci přibývání počtu synapsí a myelinizace (obr. 1).

KOGNITIVNÍ KONTROLA
Pojmem kognitivní kontrola se označuje množina procesů významných pro dosahování cílů v blízké i vzdálení budoucnosti: jde zejména o adaptivní myšlení, prožívání i akce ve vztahu k proměnám zevního prostředí. Kognitivní kontrola se objevuje v poměrně raném dětství, v dalším průběhu dětství a v adolescenci se zdokonaluje. Byť se výsledky jednotlivých studií v podrobnostech liší, závisí vývoj i míra kognitivní kontroly na činnosti řady oblastí prefrontální a temenní kůry, jakož i limbického systému (29, 30, 37). Mezi základní kontrolní funkce lze počítat například pracovní paměť nebo „přepínání“ mezi dvěma úlohami. U dospělých zátěž pracovní paměti aktivuje zevní prefrontální a temenní kůru (31). Experimenty porovnávající vysokou zátěž prostorové a slovní pracovní paměti se zátěží nízkou dokládají rostoucí aktivitu ventrální a dorzolaterální prefrontální jakož i parietální kůry ve věkových skupinách 7–12 let, 13–17 let a v dospělosti. Lze mluvit o lineárním růstu aktivity od 7. roku věku do dospělosti. V některých doménách pracovní paměti, například v útlumu odpovědi na podnět nebo v průběhu „přepínání“ jedné úlohy do druhé, byla u adolescentů v porovnání s dětským věkem zjištěna nižší aktivita horních částí zevní i vnitřní prefrontální kůry, což se interpretuje jako projev zvýšené efektivity těchto sítí. Některé oblasti prefrontální kůry tedy s věkem aktivitu zvyšují, jiné ji úměrně efektivitě sítí snižují, zatímco aktivita temenní kůry při těchto typech zátěže s věkem víceméně lineárně roste. Tradovaná představa, že za svízelemi chování doprovázejícími adolescenci, například vysokou mírou impulzivity, sklonem k nepřiměřenému riskování stojí pouhá „nezralost“ prefrontálních systémů, resp. „relativně zpomalený vývoj“, je tedy zjednodušená představa (13).
Schopnost uvažovat ve vztazích (relational reasoning) se považuje za základní složku dynamické inteligence (fluid intelligence), která vyžaduje k dosažení cíle souběžné užití většího počtu slovních nebo prostorových možností. Je podkladem komplexních kognitivních kontrolních funkcí, například monitorování vlastního výkonu nebo učení na základě zpětné vazby. V průběhu rané adolescence aktivuje positivní zpětná vazba činnost prefrontální a temenní kůry, v dospělosti však aktivuje stejné korové oblasti negativní zpětná vazba, střední adolescence je v tomto smyslu přechodnou fází (49, 50).
Z dosavadních studií plyne, že jsou systémy kognitivní kontroly pružnější a proměnlivější, než by odpovídalo jednoduché představě „zapojování on-line úměrnému věku“. Zde je možný neuronální podklad adaptibility dospívajících na sociální změny, například rychlé učení změnám v jazyce, technologiích, hudbě a módě v ošacení, stejně jako jejich adaptace po imigraci do jiného státu (17). Významná je motivace daná například přítomností věkových vrstevníků a/nebo afektivním hodnocením situace v nějakém druhu sociální situace (28). Funkční systémy čelních laloků dospívajících jsou tedy citlivé na kontext a jejich aktivitu lze zvýšit tréninkem. Možné je, že flexibilita těchto systémů daná tréninkem je vyšší než u lidí dospělých, což však bude vyžadovat ověření.
ZPRACOVÁVÁNÍ AFEKTIVNÍCH A SOCIÁLNÍCH INFORMACÍ
Zpracovávání afektivních informací se týká jak emocí a motivací, tak systémů, které vyhodnocují, jsou s to učit se o odměnách a trestech, ve vztahu k podnětu regulují základní „přibližování“ a „vzdalování“. Prožívání odměn a trestů se v průběhu adolescence odehrává zejména v sociální sféře. Pro adolescenty je významný obdiv, souhlas, odmítnutí (36, 46), přijetí jejich věkovými vrstevníky. Na odměnu odpovídá zvýšenou aktivitou ventrální striatum (nc. accumbens). Jedním z testů je v tomto směru předpověď omylu. Aktivita je v průběhu testování nejvyšší u dospívajících, u dětí a dospělých je menší (11). Významnější je však patrně funkční konektivita mezi ventrálním striatem a mediální prefrontální kůrou. Obrázky šťastných tváří aktivují ventrální striatum adolescentního mozku výrazněji než u lidí dospělých. Jestliže se dívají adolescenti staří 12–18 let, to je období označované jako střední adolescence, na obrázky ustrašených tváří, je aktivita jejich amygdaly, uzlu sítě, která zpracovává negativní afektivní podněty, opět výraznější, než je tomu u lidí dospělých (26). Sítě, jejichž uzly jsou ventrální striatum a amygdala, tedy v průběhu dospívání odpovídají intenzivněji na afektivní podněty, než je tomu v dětství a v dospělosti. Vrchol této akcentované odpovědi lidé dosahují v průběhu střední adolescence. Výsledkem jsou vyšší nároky na emoční regulaci a sebekontrolu.
Současný výzkum definoval soubor neuronální sítí metaforicky označený pojmem „sociální mozek“ (29) (obr. 2). Pojem sociálně-kognitivní vývoj označuje změnu poznávacích dovedností a znalostí, která usnadňuje pochopení sociálních situací, příkladem je mentalizace a schopnost odhadnout perspektivu (perspective-taking ability). Mentalizace je schopnost soudit na niterné duševní stavy druhých lidí, například na jejich záměry, druh přesvědčení a touhy. Mentalizace (také teorie duševních stavů, theory of mind) je v průběhu dospívání klíčová.

Základní sociální dovednosti včetně mentalizace se vyvíjejí v průběhu dětství, složitější sociálně-kognitivní dovednosti až v adolescenci. Úlohy testující mentalizaci aktivují síť, jejímiž uzly jsou kůra zadní části sulcus temporalis superior, kůra na hranicích spánkového a temenního laloku (temporoparietální funkce, TPJ), přední spánková kůra včetně kůry pólu spánkových laloků a mediální prefrontální kůra. Přední horní prefrontální kůra se aktivuje, jakmile lidé uvažují o psychologických stavech, bez ohledu na to, zda je o stavy vlastní nebo stavy druhých lidí, například vlastní maminky, zda si lidé představují druhé lidi nebo zvířata (4, 51). Nejrůznější druhy mentalizační zátěže ve všech typech experimentů aktivují dorsomediální prefrontální kůru adolescentů v porovnání s kontrolní úlohou u adolescentů více, než je tomu u lidí dospělých (obr. 3).
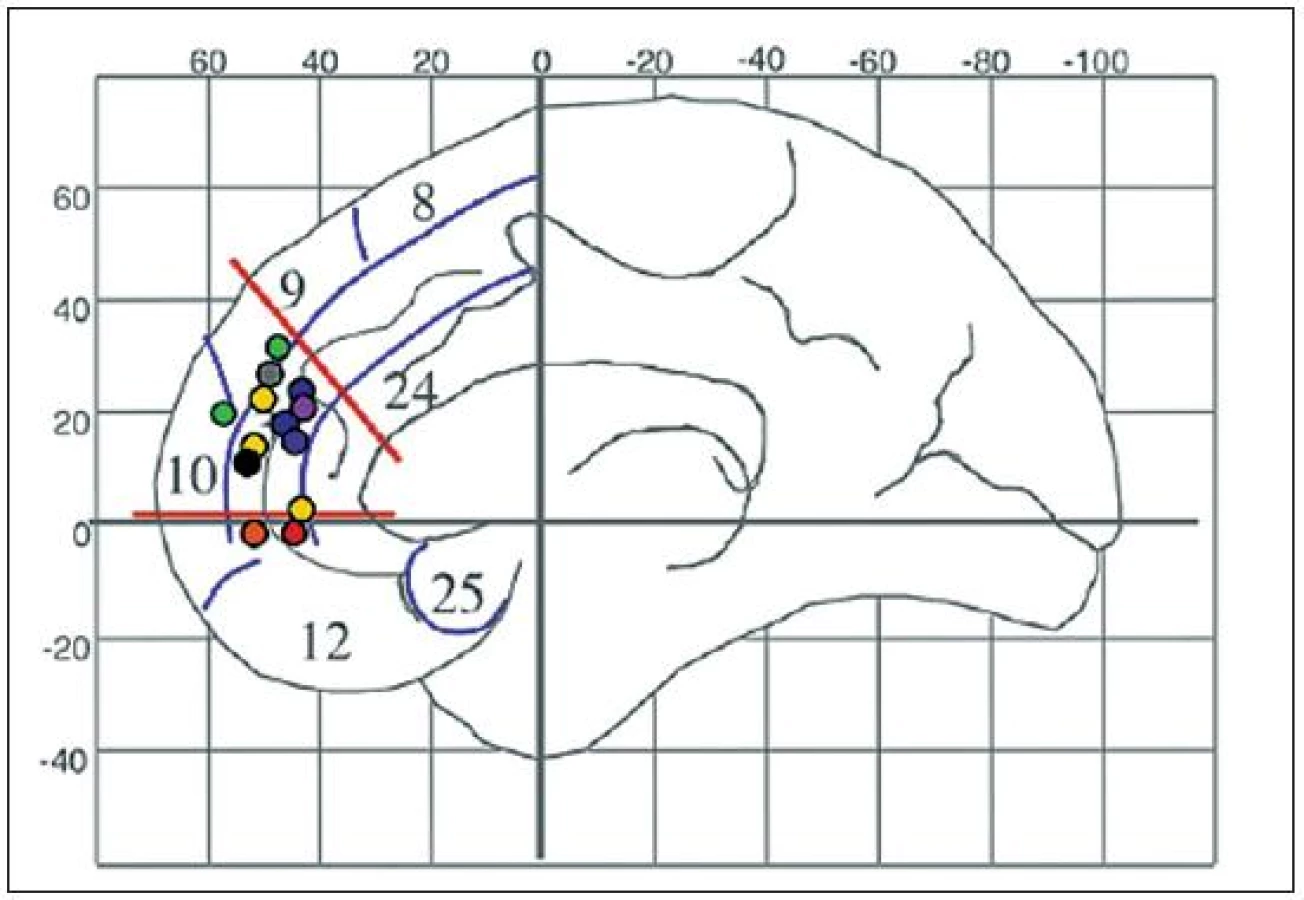
Nové studie se zaměřují na emoční aspekty mentalizace, posun aktivity mezi dětstvím a dospělostí je podobný. Burnettová et al. (10) vystavila adolescenty ve věku 10–18 let a lidi dospělé ve věku 22–32 let scénářům, z nichž plynuly sociální emoce, například pocit viny. Na rozdíl od emocí základních sociální emoce aktivují sociální mozek: dorzomediální prefrontální kůra se vysoce aktivuje u adolescentů (obr. 3 – růžové kroužky). Levostranná přední spánková kůra vykazuje opačný typ aktivity. U dospělých lidí ji vysoce aktivují sociální emoce v porovnání s emocemi základními. Relativní aktivita v průběhu sociálních aspektů mentalizace se tedy opět s věkem opět přesouvá z dorzomediální prefrontální kůry do přední temporální kůry.
Shamay-Tsoory et al. (41) rovněž rozlišuje kognitivní mentalizaci (pochopení myšlenek a úvah druhého člověka) od mentalizace afektivní (pochopení emocí druhého člověka). Adolescenti ve věku 11–16 let a lidé dospělí ve věku 24–40 let se dívali na obrázky s kognitivní a afektivní mentalizací (40). Oba druhy mentalizace aktivovaly sociální mozek. Afektivní mentalizace aktivovala ventrální mediální prefrontální kůru u adolescentů v porovnání s lidmi dospělými více než mentalizace kognitivní (obr. 3 – oranžové kroužky).
Gunther-Moor et al. (23, 24) porovnali aktivitu mozku raných adolescentů (věk 10–12 let), adolescentů ve středním období (14–16 let) a mladých dospělých lidí (19–23 let) v testu „duše v očích“ (1). V průběhu tohoto testu pozorovatelé soudí na duševní stav a emoce pouze z fotografie očí pozorované osoby. Ve všech věkových skupinách byla u pozorovatelů v porovnání s kontrolou, kterou bylo pozorování stejného podnětu a cílem odhadnout věk a pohlaví, zjištěna vyšší aktivita kůry zadní části sulcus temporalis superior. Nadto byla zjištěna ve skupině raných adolescentů zvýšená aktivita dorzomediální prefrontální kůry (obr. 3 – černé kroužky).
Dynamiku vývoje sociálního mozku v průběhu adolescence shrnuje obrázek 4. Adolescentům ve věku 12–18 let byly předváděny obrázkové scénáře záměrného jednání, v nichž se skrýval záměr a jeho důsledky nebo scénáře fyzikální kauzality (4–6). Záměrné scénáře v porovnání s fyzikální kauzalitou adolescentům aktivovaly dorzomediální prefrontální kůru více než lidem dospělým. A naopak – kůra pravostranné zadní části sulcus temporalis superior v této modalitě experimentu aktivovala více u lidí dospělých než u adolescentů. Mentalizace tedy aktivuje totožnou síť, nicméně míra aktivity se s věkem přesouvá z dorzomediální prefrontální kůry do kůry zadní části sulcus temporalis superior. Ve dvou podobných úlohách byly tázány děti a lidé dospělí, zda popsané charakterové rysy odpovídají spíše jim samotným nebo někomu jinému. Dorzomediální prefrontální kůra byla při vyvolávání informací o vlastní osobě aktivnější u dětí než u lidí dospělých.

Jednou z klíčových změn sociální interakce v průběhu dospívání bývá posun z chování orientovaného na sebe k chování orientovanému na druhé lidi. Tento přesun lze modelovat a vyšetřovat ekonomickými hrami, například hrou Konec smlouvání (Ultimatum game) a hrou Důvěra (32). Děti a raně dospívající jedinci ve věku 9–12 let chápou záměr druhých lidí hůře, v průběhu vyzrávání postupně při rozhodování berou v úvahu perspektivu druhých lidí. Úměrně tomu roste aktivita kůry TPJ (a zevní horní prefrontální kůry), zatímco aktivita přední vnitřní prefrontální kůry klesá (25). Ve hře Důvěra byli účastníci pokusu v průběhu investování druhými hráči. První hráč druhému hráči důvěřuje a svěří mu peníze. Druhý hráč se může chovat recipročně a rozdělit se s prvním hráčem rovným dílem, nebo podrazit a nechat si většinu peněz pro sebe. Rozhodnutí o případné reciprocitě vyžaduje mentalizaci. S rostoucím věkem jsou adolescenti s to lépe zaujmout perspektivu druhého jedince, což se může projevit recipročním chováním, čemuž odpovídá aktivita levostranné temporoparietální funkce a pravostranné dorzolaterální prefrontální kůry. Mladší adolescenti naproti tomu aktivovali přední vnitřní prefrontální kůru, což je uzel implicitního systémů aktivovaný zpracovávání informací cílených na sebe jakož i mentalizací. Asynchronní vývoj těchto systémů může být podkladem přesunu uvažování o sobě k uvažování o druhé osobě (50).
Souhrnně řečeno: Aktivita vnitřní prefrontální kůry v průběhu mentalizace tedy od dětství do rané dospělosti klesá. Možné výklady jsou dva: 1. mentalizace u lidí dospělých může být automatičtější, než je u dětí a dospívajících, 2. pokles aktivity může být důsledkem relativního snížení objemu šedé hmoty, jež může souviset se zánikem přebytečných synapsí (5).
Sociálně afektivní vývoj je pojem označující proměny emočních aspektů zpracovávání sociálních informací, například empatie, růstu významu, který adolescenti přikládají obdivu i přátelského chování, druhých lidí vůči sobě, stejně jako vývoj afektivních dovedností, které jsou podkladem sociální kompetence (13).
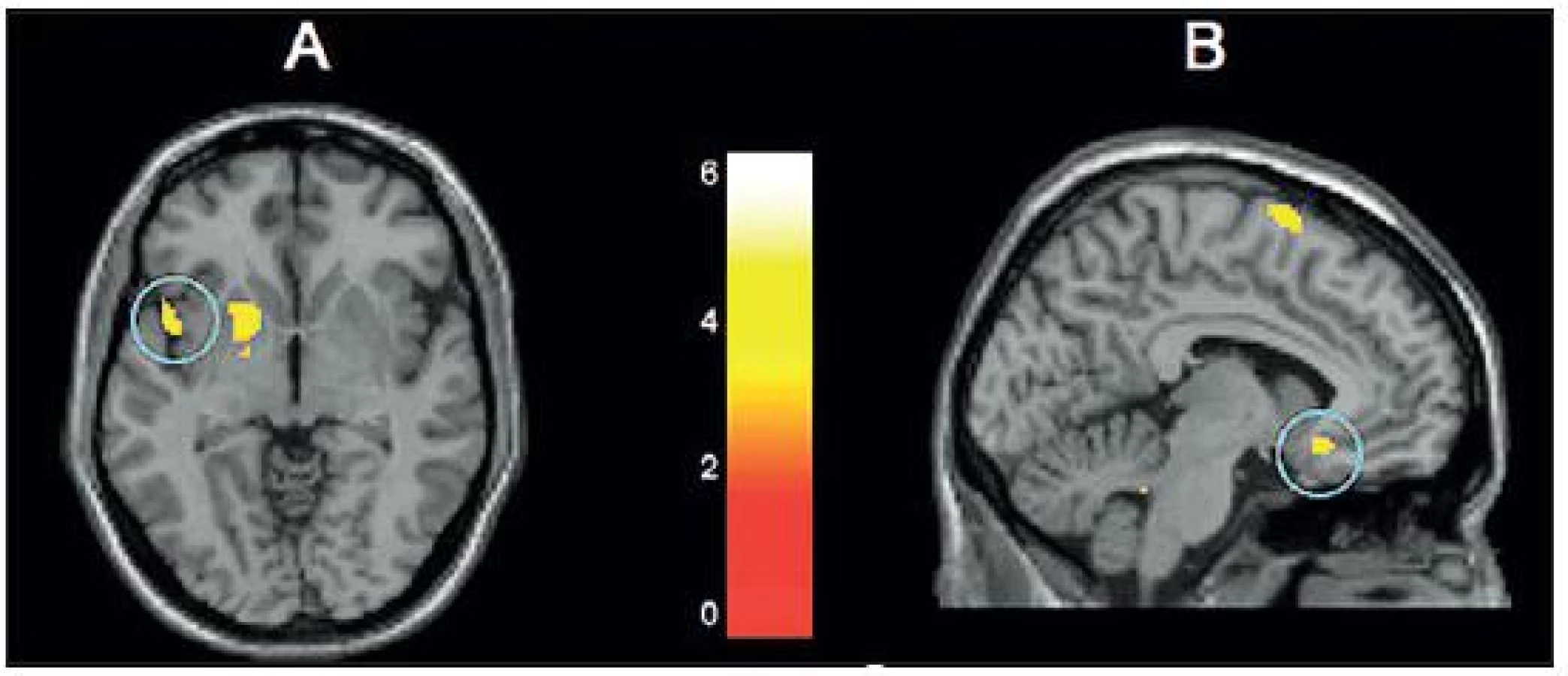
Studium empatie, sociálního přijetí i odmítnutí ukazuje rozdíly aktivity těch neuronálních sítí, které zpracovávají afekt a sociální bolest (podmíněnou například vyloučením ze skupiny) mezi dětmi, dospívajícími a lidmi dospělými, a to včetně insuly a temporálního pólu. Gunther Moor et al. (24) vyšetřovali neuronální odpověď dětí a adolescentů ve věkových skupinách 8–10 let, 12–14 let, 16–17 let jakož i dospělé lidi na sociální přijetí a odmítnutí. Jestliže účastníci pokusu dostali zpětnou vazbu, že je jejich věkoví vrstevníci mají rádi („like them“), pak se v porovnání se zpětnou vazbou tvrdícím, že je rádi nemají, zvýšila aktivace ventrální části přední cingulární kůry a striatum, a to ve všech věkových skupinách. Sociální přijetí je tedy významné ve všech věkových skupinách i v dospělosti. Sociální odmítnutí aktivovalo insulu a dorzální část přední cingulární kůry. K testování odpovědi na sociální odmítnutí, resp. vyloučení, se často užívá Cyberball test: Účastníci experimentu se stanou členy skupiny hrající počítačovou míčovou hru. Po nějaké době, na ně ostatní účastníci přestanou reagovat, „vyloučí je“. Vyloučení tohoto druhu aktivuje u dospívajících subgenuální oblast přední cingulární kůry. Jestliže mají tito adolescenti v průběhu 2 let před experimentem značné množství přátel, je aktivita jejich insulární kůry, která se v průběhu vyloučení rovněž zvyšuje, menší. Větší množství přátel tedy „imunizuje“ proti bolesti ze sociálního vyloučení (35). Stejná výzkumná skupina dokázala, že vyšší aktivita subgenuální přední cingulární kůry a vnitřní prefrontální kůry, která je ve věkové skupině 12–13 let odpovědí na experimentální sociální vyloučení předpovídá vyšší výskyt příznaků deprese v roce, který následuje po experimentu (36) (obr. 5).
Neuronální podklady hudební konformity tak známé ve skupinách adolescentů studoval Berns et al. (3). Věkový rozptyl zkoumané skupiny byl 12–17 let. Tato věková skupina hudbu nakupuje rozsáhle. Rozhodování o spotřebě a nákupu je ovlivňováno sociálně. Předpokládá se, že vkus je podmíněn popularitou dané hudby v referenční skupině. Autoři užili 15 sekundové klipy z MySpace.com. a změřili behaviorální i neuronální odpověď. Účastníci experimentu buď znali, nebo neznali míru popularity dané písničky. Čím byla populárnější, tím byla mezi pokusnými osobami oblíbenější. Mezi skóre obliby a aktivitou oblasti nc. caudatus byla zjištěna positivní korelace, tato oblast je součástí sítě centrální odměny. Tendence měnit hodnocení písničky odpovídala aktivitě přední cingulární kůry. Ta je součástí sítě orientované pozornosti, dále aktivitou přední insulární kůry, její aktivita odpovídá negativním afektivním stavům. Citlivost k míře popularity odpovídala snižování aktivity kůry středního spánkového závitu, což odpovídá nižší úrovni zpracovávání sémantické informace. Autoři předpokládají, že hlavním faktorem, který ovlivňuje spotřebitelskou volbu, je míra úzkosti pramenící z odlišnosti vlastní preference a preference druhých lidí, což vede k přesmyku volby tak, aby odpovídala vkusu, resp. volbě referenční skupiny.
Je sice zřejmé, že počátek puberty orientuje sociální vztahy směrem k věkovým vrstevníkům a posunuje rovnováhu zpracovávání afektu porovnávajícího odměnu s hrozbou ve prospěch odměny. Způsob, jakým mění jednotlivé hormony stavbu a činnost funkčních systémů mozku odpovídajících za motivaci a emoční zpracovávání směrem k vyhledávání senzací, však zůstává nejasný.
Podobně nejasný je vztah puberty a věku per se. Tradovaný model vývoje adolescentního mozku kladoucí důraz na relativně opožděné vyzrávání prefrontálních funkčních systémů, které má vysvětlovat některé rizikové stránky chování dospívajících, se mění. Současný model předpokládá asociaci začátku puberty se změnami stavby a činnosti těch funkčních systémů mozku, které zpracovávají sociální a afektivní informace. Předpokládá se, že rychlý vzestup hladiny pohlavních hormonů ovlivňuje zejména limbický systém, zejména jde o proměny stavby a činnosti ventrálního striata (nc. accumbens) a amygdaly, včetně jejich korových projekcí. Puberta navodí zvýšené zpracovávání podnětů, které vedou k vyhledávání senzací, nových událostí (novelty seeking), zvýší zpracovávání informací, které mají vztah k poloze na sociálním žebříčku, například zvýší vnímavost vůči pozornosti a obdivu věkových vrstevníků. Změny tohoto druhu se v průběhu adolescence stupňují, vrcholí ve středním dospívání, přetrvávají několik let. Vývoj může být hodnotově i sociálně pozitivní, stejně jako negativní, směřující k potenciálně kriminálnímu chování nebo zneužívání alkoholu a drog nebo k depresi (obr. 3).
Genderové rozdíly popsali Giedd et al. (20). Největší objem dosahuje šedá hmota čelních a temenních laloků v 11 letech u dívek, ve 12 letech u chlapců, což přibližně odpovídá začátku pubertální činnosti gonád obou pohlaví. Mezi růstem hladiny pohlavních hormonů a růstem objemu šedé hmoty tedy může být souvislost. Jiné studie zjistily postupný vývoj pohlavního dimorfismu mozku v průběhu adolescence. Objem amygdaly roste u chlapců, objem hipokampu u dívek. Neufang et al. (38) zkoumal vztah mezi objemem šedé hmoty, pohlavím a známkami puberty v souboru dětí a dospívajících ve věkovém rozmezí 8–15 let. Bez ohledu na pohlaví zjistil pozitivní vztah mezi znaky puberty a objemem šedé hmoty amygdaly a negativní vztah mezi těmito známkami a objemem hipokampu. U dívek skupina zjistila positivní vztah mezi hladinou estrogenu a objemem limbické šedé hmoty. U chlapců byl zjištěn negativní vztah mezi hladinou testosteronu a objemem temenní šedé hmoty. Bramen et al. (8) vyšetřoval pohlavní rozdíly ve vztahu ke stavbě mozku u 80 adolescentních dívek a chlapců, kteří byli „srovnáni“ co do pohlavní zralosti, nikoliv co do věku. Skupina dokázala signifikantní interakce mezi stupněm pohlavní zralosti, pohlavím a korovými oblastmi s vysokou hustotou receptorů pohlavních hormonů. Pohlavní rozdíly byly větší v pravém hipokampu, amygdale oboustranně a korové šedé hmotě u pohlavně zralejších adolescentů. Větší objem šedé hmoty byl zjištěn v mediálních oblastech spánkových laloků u pohlavně zralejších chlapců, u pohlavně zralejších dívek byl objem zdejší šedé hmoty menší (6).
Extroverze a neuroticismus (neboli negativní emocio-nalita) jsou dva rozměry Velké pětky, pětirozměrového modelu osobnosti (12). U ženského pohlaví predisponují k chronickým bolestivým stavům a afektivním poruchám více než u pohlaví mužského. Blankestein et al (7) uvedli do vztahu profil osobnosti a stavbu mozku u dospívajících dívek a chlapců. Míra extroverze korelovala u dívek záporně s objemem šedé hmoty středního frontálního závitu. Míra neuroticismu u dospívajících dívek korelovala pozitivně s tloušťkou mozkové kůry a objemem šedé hmoty přední části cingulárního závitu. U chlapců byl trend opačný.
Dosud poměrně malou pozornost věnoval výzkum strukturální a funkční konektivitě a její účinnosti, byť její rozdíly mohou v nějaké míře vysvětlovat rozdíly v chování, poznávacích funkcích i afektivitě. Klidová funkční konektivita (rsfMRI, resting state fMRI) odpovídá konektivitě mozku, jenž je v „klidu“ bez jakékoliv zevní zátěže. Hlavní obvody kognitivní kontroly jsou vyvinuté již na začátku adolescence a v jejím průběhu vyzrávají. Spojení na krátké vzdálenosti v průběhu času slábnou, naproti tomu spojení na vzdálenosti dlouhé, o nichž se předpokládá, že jsou významné pro integraci činnosti jednotlivých funkčních systémů, posilují. Zpočátku slabší konektivita integrující obvody propojující systémy na dlouhé vzdálenosti může být podkladem varia-bilního výkonu dospívajících lidí při vysokých nárocích na pozornost a rozhodování – jejich schopnost integrovat je méně automatická než u lidí dospělých – na druhé straně může být podkladem schopnost odpovídat adaptivně a tvořivě, nerutinně. Součástí této konektivity je také implicitní (default) systém, jehož aktivitu lze považovat za jeden ze základních aspektů sebe-uvědomování. I tento systém se vyvíjí od dětství do dospělosti (33).
Konektivitu úměrnou zátěži (task dependent connectivity) lze zkoumat dvěma způsoby. Jedním z nich je analýza psychofyziologické interakce (PPI, psychophysiological interaction). To je statistická technika založená na lineární regresi, výsledky jsou nezávislé na výsledcích zjištěných konvenční analýzou: Jestliže aktivita mozkové oblasti A předpovídá aktivitu mozkové oblasti B, pak statistická mohutnost předpovědi odpovídá vlivu, který má oblast A na oblast B. Jestliže se mohutnost předpovědi proměňuje v tom psychologickém kontextu, v němž se měří fyziologická aktivita, pak se jev považuje za důkaz psychofyziologické interakce (15). Analýza tohoto druhu dokázala ve výše zmíněném experimentu cíleném na testování mentalizace vyšší míru funkční konektivity v průběhu sociálních než v průběhu základních emocí jak u dospívajících, tak dospělých. Předpokládá se vyzrávání a efektivnější specializace podsystémů mentalizační sítě.
Burnettová a Blakemoreová (10) užily analýzu psychofysiologické interakce v souboru adolescentů starých 11–18 let a v souboru mladých dospělých lidí ve věku 22–32 let. Pokusné osoby byly vyšetřovány fMR v době, kdy uvažovaly o scénářích, z nichž plynuly základní emoce, například hnus a strach a emoce sociální, například pocit viny nebo znepokojení. Sociální emoce vyžadují mentalizaci. Jak u dospívajících, tak u lidí dospělých aktivovaly sociální emoce ve větší míře funkční konektivitu předních částí vnitřní prefrontální kůry jednak s kůrou zadní části s. temporalis superior v místě, kde hraničí s temporoparietální junkcí a kromě toho s přední spánkovou kůrou. Složky systému mentalizace jsou tedy v průběhu sociální emoční zátěže ve vzájemné interakci. Mezi dospívajícími a dospělými však byl zjištěn rozdíl: U dospívajících byla v průběhu socio-emoční zátěže v porovnání se zátěží danou základními emocemi doložena mohutnější konektivita mezi přední vnitřní prefrontální kůrou a kůrou zadní části sulcus temporalis superior, než tomu bylo u lidí dospělých. Systém tedy s věkem vyzrává.
Druhým způsobem studia konektivity je například dynamické kauzální modelování (DCM, dynamic causal modelling (16), které zjišťuje jaký směr má vliv jedné mozkové oblasti na druhou. Studie tohoto druhu doložila u dětí a dospívajících slabší modulační vliv „shora – dolů“ (top-down), který mají čelní oblasti na „nižší“ oblasti mozku, než je tomu u lidí dospělých (27). U dětí a dospívajících jsou tedy „nižší“ oblasti mozku pod menší mírou kontroly funkčními systémy čelních laloků (30), než je tomu u lidí dospělých.
Buněčné podklady popsaných změn aktivity nejsou známy. Předpokládá se, že za nimi jsou proměny stavby a funkce synapsí. Úvaha říká, že relativní „nadbytek“ synapsí v prefrontální kůře a dalších mozkových oblastech v dětství a rané adolescenci je důvodem menší výkonnosti daných funkčních systémů. Ty ji v plnější podobě dosáhnout synaptickou přestavbou sice ovlivněnou evolučně geneticky nicméně danou vlivy prostředí. Relativně nadbytečný počet synapsí může být důvodem nízkému poměru signál/šum. Poté, co nadbytečné synapse zmizí, je menší počet zbylých synapsí výkonnější. Skutečnost však bude složitější. Různé mozkové oblasti se vyvíjejí různým tempem, jejich stavební a funkční proměny nejsou paralelní. Stejné mozkové oblasti v průběhu některých úloh aktivitu zvyšují, v průběhu dalších úloh aktivitu snižují. Při zátěži nějakou úlohou nemusí být nutně aktivní všechny místní synapse. Vztah mezi aktivitou synapsí a krevním průtokem nemusí být nutně lineární a může být v různých oblastech mozku a různých lidí při různých úlohách různý.
SYSTÉM MOTIVACE A ODMĚNY
Charakteristické adolescentní chování, jehož složkami je vydatně zvýšený podíl interakcí s věkovými vrstevníky, zvýšená míra rizikového chování, vyhledávání senzací a nových událostí, se objevuje nejen u lidí. Zřejmě jde o selektovaný druh chování podněcující vyhledávání nových území, pohlavních partnerů a zdrojů potravy (45). Interakce s věkovými vrstevníky začne ovlivňovat rozhodování a chování ve větší míře, než je tomu u dospělých lidí. Interakce s věkovými vrstevníky je pro adolescenty druhem sociální odměny. Dospívající riskují více než dospělí (47). Růst míry rizikového chování je spjat s motivací zkoušet nové podněty, které mohou být zdrojem odměny. Motivace tohoto typu je jedním z podkladů testování tabáku, alkoholu a drog, z nichž může vyplynout zneužívání a závislost.
Neuronálním podkladem získávání odměny a motivace je vývojově starý systém, jehož osou jsou nc. accumbens (ventrální striatum) a dopaminergní nc. tegmentalis ventralis v horním kmeni. Součástí jsou dopaminergní projekce směřující z nc. tegmentalis ventralis do amygdaly, hipokampu a prefrontální kůry společně s rovněž dopaminergním nigrostriatálním systémem, jenž propojuje s. nigra a dorzální striatum. Funkční význam jednotlivých složek systému je předmětem probíhající diskuze (2). V průběhu adolescence prodělává tento systém u zvířat, například experimentálních potkanů, stejně jako u lidí rozsáhlou, přechodnou, nelineární a dynamickou přestavbu. Například přibývají projekce z prefrontální kůry do nc. accumbens. Roste hustota dopaminergních receptorů v dorzálním striatu, v pozdní adolescenci je o 40 % vyšší než v mladším věku a v dospělosti (9), v dospělosti počet dopaminergních synapsí této oblasti opět klesá. V nc. accumbens je pokles podílu dopaminergních synapsí nižší. Některé studie uvádějí, že neklesá (14).
ADRESA PRO KORESPONDENCI:
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Thomayerova nemocnice
Vídeňská 800, 140 59 Praha 4 – Krč
e-mail: frantisek.koukolik@ftn.cz
Sources
1. Baron-Cohen S, Jollife T, Mortimore C. Another advanced test of theory of mind: evidence from very high functioning adults with autism or Asperger syndrome. J Child Psychol Psychiatry 1997; 38 : 813–822.
2. Berridge KC, Kringelbach ML. Affective neuroscience of pleasure: reward in humans and animals. Psychopharmacology (Berl) 2008; 199 : 457–480.
3. Berns GS, Capra CM, Moore S, et al. Neural mechanisms of the influence of popularity on adolescent ratings of music. NeuroImage 2010; 49 : 2687–2696.
4. Blakemore SJ, den Ouden H, Choudhury S, et al. Adolescent development of the neural circuitry for thinking about intentions. Soc Cogn Affect Neurosci 2007; 2 : 130–139.
5. Blakemore SJ. The social brain in adolescence. Nature Rev Neurosci 2008; 9 : 267–277.
6. Blakemore SJ. Imaging brain development: the adolescent brain. NeuroImage 2012; 2 : 397–406.
7. Blankestein U, Chen JYW, Mincic AM, et al. The complex mind of teenagers: neuroanatomy of personality differs between sexes. Neuropsychologia 2009; 47 : 599–603.
8. Bramen JE, Hranilovich JA, Dahl RE, et al. Puberty influences medial temporal lobe and cortical gray matter maturation differently in boys than girls matched for sexual maturity. Cereb Cortex 2011; 21 : 636–646.
9. Brenhouse HC, Andersen SL. Delayed extinction and stronger reinstatement of cocaine conditioned place preference in adolescent rats, compared to adults. Behav Neurosci 2008; 122 : 460–465.
10. Burnett S, Blakemore SJ. Functional connectivity during a social emotion task in adolescents and in adults. Eur J Neurosci 2009; 29 : 1294–1301.
11. Cohen JR, Asarnow RF, Sabb FW, et al. A unique adolescent response to reward prediction errors. Nat Neurosci 2010; 13 : 669–671.
12. Costa PT, McCrae RR. The revised NEO personality Inventory (NEO PI-R) professional manual. Odessa, FL: Psychological Assessment Resources 1992.
13. Crone EA, Dahl RE. Understanding adolescence as a period of social-affective engagement and goal flexibility. Nat Neurosci 2012; 13 : 637–650.
14. Doremus-Fitzwater TL, Varlinskaya EI, Spear LP, et al. Motivational systems in adolescence: possible implications for age differences in substance abuse and other risk-taking behaviors. Brain Cogn 2010; 72(1): 114. Published online 2009 September 16. doi:10.1016/j.bandc.2009.08.008
15. Friston KJ, Buechel C, Fink R, et al. Psychophysiological and modulatory interactions in neuroimaging. NeuroImage 1997; 6 : 218–229.
16. Friston KJ, Harrison L, Penny W. Dynamic causal modelling. NeuroImage 2003; 19 : 1273–1302.
17. Fuligni AJ. Adolescents from immigrant families. In: McLoyd V, Steinberg L. (eds.) Studying minority adolescents: Conceptual, theoretical, and methodological issues. Mahwah, New Jersey: Lawrence Erlbaum Assoc 1998 : 127–143.
18. Giedd JN, Blumenthal J, Jeffries NO, et al. Brain development during childhood and adolescence: a longitudinal MRI study. Nat Neurosci 1999; 2 : 861–863.
19. Giedd JN. Structural magnetic resonance imaging of the adolescent brain. Ann NY Acad Sci 2004; 1021 77–85.
20. Giedd JN, Raznahan A, Mills KL, et al. Review: magnetic resonance imaging of male/female differences in human adolescent brain anatomy. Biol Sex Differ 2012; 3(19): 1–9.
21. Giedd JN, Keshavan M, Paus T. Why do many psychiatric disorders emerge during adolescence? Nat Rev Neurosci 2008; 9 : 947–958.
22. Gogtay N, Giedd JN, Lusk L, et al. Dynamic mapping of human cortical development during childhood through early adulthood. Proc Natl Acad Sci USA 2004; 101 : 8174–8179.
23. Gunther Moor B, Op de Macks ZA, Guroglu B, et al. Neurodevelopmental changes of reading the mind in the eyes. Soc Cogn Affect Neurosci 2012; 7(1): 44–52. Published online 2011 April 22. doi:10.1093/scan/nsr020.
24. Gunther Moor B, van Leijenhorst L, Rombouts SA, et al. Do you like me? Neural correlates of social evaluation and developmental trajectories. Soc Neurosci 2010; 5 : 461–482.
25. Guroglu B, van den Bos W, van Dijk E, et al. Dissociable brain networks involved in development of fairness considerations: understanding intentionality behind unfairness. Neuroimage 2011; 57 : 634–641.
26. Guyer AE, Monk AS, McClure-Tone EB, et al. A developmental examination of amygdala response to facial expressions. J Cogn Neurosci 2008; 20 : 1565–1582.
27. Hwang K, Velanova K, Luna B. Strengthening of top–down frontal cognitive control networks underlying the development of inhibitory control: a functional magnetic resonance imaging effective connectivity study. J Neurosci 2010; 30 : 15535–15545.
28. Johnson MH, Grossmann T, Cohen Kadosh K. Mapping functional brain development: building a social brain through interactive specialization. Dev Psychol 2009; 45 : 151–159.
29. Koukolík F. Základy sociální, kognitivní a afektivní neurovědy XI. Sociální rozhodování. Prakt. Lék. 2011; 91 : 632–638.
30. Koukolík F. Řídící funkce čelních laloků. In: Lidský mozek. Funkční systémy. Norma a poruchy. 3. Přepracované a doplněné vydání. Praha: Galén 2012; 353–378.
31. Koukolík F. Pracovní paměť. In: Lidský mozek. Funkční systémy. Norma a poruchy. 3. Přepracované a doplněné vydání. Praha: Galén 2012; 117–123.
32. Koukolík F. Zvíře politické. Praha: Galén 2012.
33. Koukolík F. Já. O mozku, vědomí a sebeuvědomování. Praha: Karolinum 2013.
34. Lenroot RK, Giedd, JN. Brain development in children and adolescents: insights from anatomical magnetic resonance imaging. Neurosci Biobehav Rev 2006; 30 : 718–729.
35. Masten CL, Eisenberger NI, Borofssky L, et al. Neural correlates of social exclusion during adolescence: understanding the distress of peer rejection. Soc Cogn Affect Neurosci 2009; 4 : 143–157.
36.Masten CL, Eisenberger NI, Borofssky L, et al. Subgenual anterior cingulate responses to peer rejection: a marker of adolescents’ risk for depression. Dev Psychopathol 2011; 23 : 283–292.
37. Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annu Rev Neurosci 2001; 14 : 167–202.
38. Neufang S, Specht, Hausmann M, et al. Sex differences and the impact of steroid hormones on the developing human brain. Cereb Cortex 2009; 19 : 464–473.
39. Petanjek Z, Judaš M, Šimic G, et al. Extraordinary neoteny of synaptic spines in the human prefrontal cortex. Proc Natl Acad Sci USA 2011; 32 : 13281–13286.
40. Sebastian CL, Fontaine NMG, Bird G, et al. Neural processing associated with the cognitive and affective theory of mind in adolescent and adults. Soc Cogn Affect Neurosci 2012; 7 : 53–63.
41. Shamay-Tsoory SG, Harari H, Aharon-Peretz J, et al. The role of the orbitofrontal cortex in affective theory of mind deficits in criminal offenders with psychopathic tendecies. Cortex 2010; 46 : 668–677.
42. Shaw P, Kabani NJ, Exkstrand K, et al. Neurodevelopmental trajectories of the human cerebral cortex. J. Neurosci 2008; 28 : 3586–3595.
43. Schlegel A, Barry H. Adolescence: An anthropological inquiry. Free Press, 1991.
44. Sisk CL, Zehr JL. Pubertal hormones organize the adolescent brain and behavior. Front Neuroendocrinol 2005; 26 : 163–174.
45. Spear LP. The developing brain and adolescent-typical behavior patterns: An evolutionary approach. In: Walker E, Romer D. (eds). Adolescent Psychopathology and the Developing Brain: Integrating Brain and Prevention Science. New York: Oxford University Press 2007; 9–30.
46. Spear LP. Heightened stress responsivity and emotional reactivity during pubertal maturation: implications for psychopathology. Dev Psychopathol 2009; 21 : 87–97.
47. Steinberg L. A social neuroscience perspective on adolescent risk-taking. Dev Rev 2008; 28 : 76–106.
48. Steinberg, L. Adolescence, 9th Edition. New York: McGraw-Hill Higher Education 2010.
49. Van den Bos W, Guroglu B, van den Bulk BG, et al. Better than expected or as bad as you thought? The neurocognitive development of probabilistic feedback processing. Front Hum Neurosci 2009; 3 : 52 [on-line]. Dostupné z: http://www.frontiersin.org/human_neuroscience/10.3389/neuro.09.052.2009/full.
50. Van den Bos W, van Dijk E, Westeberg M, et al. Changing brains, changing perspectives: the neurocognitive development of reciprocity. Psychol Sci 2011; 22 : 60–70.
51. Van Overwalle F. Social cognition and the brain: a meta-analysis. Hum Brain Mapp 2009; 30 : 829–858.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2013 Issue 6
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
Most read in this issue
- Neurobiologie dospívání
- Nová doporučení v hodnocení EKG u sportovců: „Seattle“ kritéria
- Kognitívna rehabilitácia u pacientov s miernou kognitívnou poruchou a demenciou v podmienkach denného stacionára – pilotná štúdia
- Násilí ve zdravotnictví