
Araucan and Caupolican Integrate Intrinsic and Signalling Inputs for the Acquisition by Muscle Progenitors of the Lateral Transverse Fate
A central issue of myogenesis is the acquisition of identity by individual muscles. In Drosophila, at the time muscle progenitors are singled out, they already express unique combinations of muscle identity genes. This muscle code results from the integration of positional and temporal signalling inputs. Here we identify, by means of loss-of-function and ectopic expression approaches, the Iroquois Complex homeobox genes araucan and caupolican as novel muscle identity genes that confer lateral transverse muscle identity. The acquisition of this fate requires that Araucan/Caupolican repress other muscle identity genes such as slouch and vestigial. In addition, we show that Caupolican-dependent slouch expression depends on the activation state of the Ras/Mitogen Activated Protein Kinase cascade. This provides a comprehensive insight into the way Iroquois genes integrate in muscle progenitors, signalling inputs that modulate gene expression and protein activity.
Published in the journal:
Araucan and Caupolican Integrate Intrinsic and Signalling Inputs for the Acquisition by Muscle Progenitors of the Lateral Transverse Fate. PLoS Genet 7(7): e32767. doi:10.1371/journal.pgen.1002186
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002186
Summary
A central issue of myogenesis is the acquisition of identity by individual muscles. In Drosophila, at the time muscle progenitors are singled out, they already express unique combinations of muscle identity genes. This muscle code results from the integration of positional and temporal signalling inputs. Here we identify, by means of loss-of-function and ectopic expression approaches, the Iroquois Complex homeobox genes araucan and caupolican as novel muscle identity genes that confer lateral transverse muscle identity. The acquisition of this fate requires that Araucan/Caupolican repress other muscle identity genes such as slouch and vestigial. In addition, we show that Caupolican-dependent slouch expression depends on the activation state of the Ras/Mitogen Activated Protein Kinase cascade. This provides a comprehensive insight into the way Iroquois genes integrate in muscle progenitors, signalling inputs that modulate gene expression and protein activity.
Introduction
In Drosophila as in vertebrates the proper function of the muscular system relies on the generation of a stereotyped pattern of discrete muscles and their intimate connection with the nervous system, which together control the adequate release of contraction power to fulfil the functional requirements of the organism. The formation of a muscle pattern is therefore of great importance and consequently many efforts have been devoted to solve the central problem of the acquisition of muscle identity. The embryonic Drosophila muscle pattern comprises thirty elements in each abdominal hemisegment (Figure 1G). Each muscle is a syncytial fibre whose unique characteristics, i.e., position, size, attachment to tendon cells, innervation and pattern of gene expression allow its unambiguous identification [1], [2]. Muscle specification is a stepwise process that ensures the local singling out of a population of myoblasts, the founder myoblasts, each of them containing the necessary information to give rise to a unique muscle. The origin of founder myoblasts can be traced to late embryonic stage 10 when groups of mesodermal cells (the promuscular clusters) start expressing the proneural gene lethal of scute and acquire myogenic competence [3]. Opposing activities of Notch and Receptor Tyrosine Kinase signalling pathways ensure that only one cell in the cluster will segregate as a muscle progenitor [4]. This will divide asymmetrically to generate two sibling founder myoblasts or a founder myoblast and an adult muscle precursor [3], [5], [6]. The unselected cells of the promuscular clusters, by activation of the Notch signalling pathway, will initiate the expression of the transcriptional regulator Myoblasts incompetent (also called Gleeful and Lame duck) and become fusion competent myoblasts that by fusing to founders will give rise to multinucleated fibres [7]–[9]. Regarding muscle identity, each progenitor and founder exhibits a specific code of gene expression that confers to muscles their unique characteristics. The components of these codes are accordingly named muscle identity genes (reviewed in [2], [10], [11]). The identity code is transmitted to all the nuclei in the syncytium through the process of myoblast fusion [12]. According to their patterns of expression muscle identity genes can be grouped into three categories. Type I includes genes expressed by progenitors and whose expression is maintained in sibling founders and muscles. Examples are apterous, ladybird (lb) and Pox meso (Poxm) [13]–[15]. Type II identity genes are expressed in progenitors but differentially regulated in sibling founders, being lost from one of them and the corresponding muscles. Examples are Krüppel (Kr), even-skipped (eve), collier and slouch (slou) [3], [4], [16]–[18]. And finally type III refers to genes expressed by progenitors and founders of muscles sharing common characteristics. vestigial (vg), expressed by all internal muscles, is the only known member of this class [12], [19]. Regarding the onset of their expression a few muscle identity genes, such as Kr, eve and collier, are already expressed in the promuscular cluster, before the segregation of muscle progenitors [4], [16], [18], [20] whereas other genes, like Connectin (Con), initiate their expression in already segregated progenitors [4], [16], [18], [20].

In this study we identify araucan (ara) and caupolican (caup), two members of the Iroquois gene complex (Iro-C), as novel type III muscle identity genes. The Iro-C genes encode homeoproteins conserved throughout the animal kingdom. They are organized in genomic clusters of three paralogous genes, one in the case of Drosophila and usually two in most vertebrates [21]. They participate in a wide variety of developmental processes, mainly related to the specification and patterning of diverse territories of the body, including the lateral mesonotum and dorsal cephalic region of Drosophila, the neural ectoderm of Xenopus and cranial placode derivatives of zebrafish [22]–[30]. Here we show by means of genetic approaches that ara and caup function redundantly in the specification of the lateral transverse (LT) muscles, since in the absence of both genes LT1–4 muscles loose their LT fates and acquire those of other muscles.
At present there is compelling evidence that muscle progenitors can integrate positional and temporal signalling inputs. This promotes the expression of unique combinations of muscle identity genes, which confers on them their ultimate fate [14]–[18], [31], [32]. There has been extensive analysis on the regulation of some of these genes, such as eve and collier [4], [33], [34], which has allowed to propose candidate cis-regulatory modules for founder muscle specific expression [35]. However, very little is known about how progenitors integrate the activity of the transcription factors encoded by these genes, about the identity of their direct targets (save in the cases of Kr and Lb [36]–[38]), and of their hierarchical relationships and their putative post-transcriptional regulation. In this report we have focused on these issues in relation to the function of the ara/caup identity genes. We demonstrate that the implementation of the lateral transverse muscle fate requires the repression mediated by Ara/Caup of the muscle identity genes slou and vg, to avoid reiteration of other muscle fates regulated by these transcription factors. In addition, we identify slou as a potential direct target of Ara/Caup. Furthermore, our tissue culture and in vivo experiments show that the repression of slou by Ara/Caup in LT precursors requires the activity of the Ras/Mitogen Activated Protein Kinase (Ras/MAPK) pathway to be kept at a very low level, since otherwise Caup is converted from a repressor to an activator of slou. This is to our knowledge the first evidence of the interplay between the Receptor Tyrosine Kinase signalling pathways and the activity of a muscle identity transcription factor. Therefore, during Drosophila embryogenesis, and for the acquisition of the lateral transverse muscle fate, the homeoproteins Ara and Caup appear to act at a nodal point in muscle progenitors, as they integrate positional and temporal signalling inputs that modulate their activity on subordinate identity genes.
Results
Expression of Iro-C genes during muscle development
The patterns of expression of ara and caup in the embryonic ectoderm have been previously reported [39], [40]. In this work we focus on the embryonic ara and caup mesodermal expression. In situ hybridization showed that here both genes were similarly expressed (Figure 1 and results not shown). At early stage 11 caup (and ara) transcripts and proteins are detected in groups of cells of the presumptive visceral trunk mesoderm (Figure 1A-1A′″, the available anti-Caup antibody recognises both Ara and Caup proteins). By mid stage 11 they are expressed at the same dorso-ventral level in the visceral mesoderm and in the dorsolateral ectoderm (Figure 1B-1B′″). Expression in the visceral mesoderm declined at late stage 11 when it became detectable in groups of cells of the somatic mesoderm (promuscular clusters [3], Figure 1C and 1C′), from where a subset of muscle progenitors (P) still expressing ara/caup, will segregate slightly later (stage 12, Figure 1D). Expression was maintained in sibling founder myoblasts (Fs in Figure 1D′) derived from ara-caup-expressing progenitors and in the muscles they give rise to (Figure 1E-1E″), namely LT1–4, dorsal transverse 1 (DT1) and segment border muscle (SBM) (Figure 1F and 1G). The expression in the somatic mesoderm of the third member of the Iroquois complex, mirror [41] did not overlap with that of ara-caup (not shown).
The early expression of ara/caup in all lateral muscles with vertical orientation, suggested a possible role as muscle identity genes. Therefore, we compared their expression with that of several muscle identity genes. For the LT1–4 muscles, ara/caup were co-expressed with Kr in the promuscular clusters from which progenitors PLT1/LT2 and PLT3/LT4 are singled out (Figure 2A). ara/caup expression was maintained at high levels in both progenitors that also express Kr (Figure 2B). Whereas Kr expression decayed in founders LT1 and LT3 before the onset of myoblast fusion and in LT2 and LT4 muscles from stage 15 onwards [18], expression of ara/caup was maintained in the four founders (Figure 2D and 2E). These also expressed Con, co-expression that was maintained in the mature LT1–4 muscles (Figure 1G). In the case of muscle DT1, the onset of ara/caup expression coincided with that of Con and slou in the progenitor of DT1 and dorsal oblique 3 (DO3) muscles (Figure 2C) and it appeared to be maintained in DT1 founder (Figure 2E) and mature muscle at low levels (Figure 1G). Finally, ara/caup co-expressed with lb in the SBM founder (Figure 2D), but were not be detected in the lb-expressing progenitor and promuscular cluster. In summary, different muscle lineages expressed ara/caup at different steps of the myogenic programme (Figure 2F). In the LT1–4 case ara/caup and Kr were detected at the earliest lineage stage, that is in promuscular clusters, preceding Con expression in progenitors (not shown); in the DT1/DO3 lineage ara/caup and slou were first detected in the already singled out DT1/DO3 progenitor and in the SBM ara/caup expression was first detected in the SBM founder after lb expression.

Ara and Caup are required for specification of lateral transverse muscles
During imaginal development Ara and Caup can functionally substitute each other in all territories where their function has been investigated [22], [23], [28]. Thus, to analyse their role in embryonic myogenesis and evaluate the possible contribution of mirror to any phenotype we might find, we used three deficiencies: Df(3L)iroDFM3, which removes both ara and caup, (and probably affects mirror regulation, [23], [28]), Df(3L)iroEGP6, which removes ara and caup without affecting mirror and its regulatory region, and Df(3L)iroEGP5, which only removes mirror [42]. Whereas Df(3L)iroEGP5 embryos did not show any detectable phenotype in the lateral region (not shown), a distortion of the lateral larval muscle pattern (visualised with antibody MAC141 to Tropomyosin) was found in both Df(3L)iroDFM3 and Df(3L)iroEGP6 embryos (Figure 3A–3C). In more than 95% of cases muscles with LT morphology were absent (Figure 3E). Instead, some fibres with abnormal orientation appeared in the lateral and ventral regions, but never inserted at the LT attachment sites (asterisks in Figure 3B and 3C). The loss of LT muscles was further verified by loss of expression of the specific LT muscle marker CG13424, recently renamed lateral muscles scarcer (lms) [43] at stage 15 and the absence of Con expression in the lateral somatic mesoderm (Figure S1). Both DT1 and SBM fibres developed with normal morphologies (Figure 3A–3C and Figure S1). To examine the individual contribution of ara and caup to the phenotype we resorted to embryos mutant for only one of these genes (ara in ararF209, [28], or caup in iroEGPΔ1, [42]). The larval muscle pattern was normal in both mutants (not shown). Thus, similarly to imaginal development, ara and caup appear to play redundant roles during embryonic myogenesis.
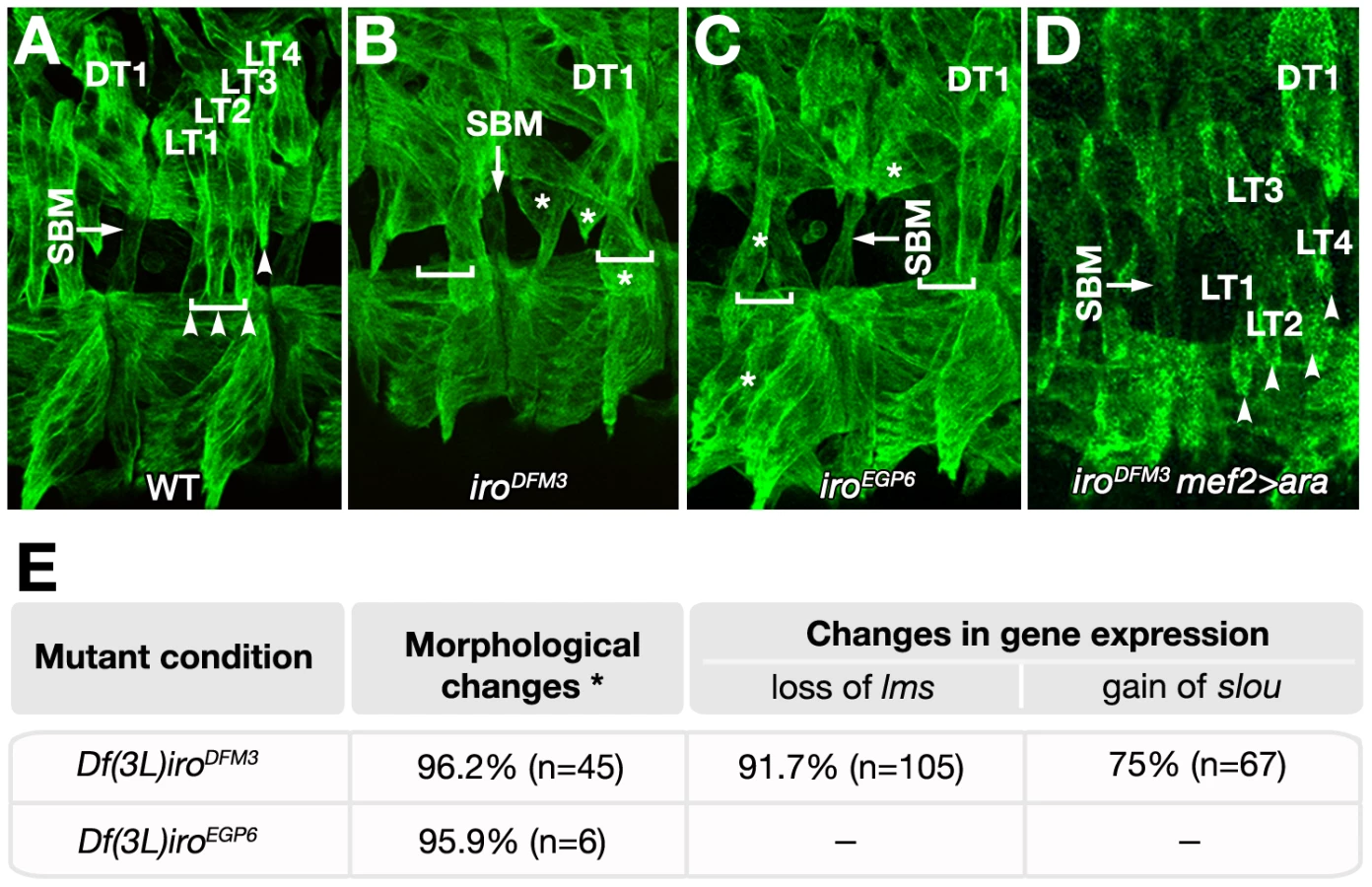
The absence of muscles with LT morphology in ara/caup mutants could be due to a failure of otherwise well specified muscles to find the right insertion to tendon cells, due to ectodermal requirement of Iro-C genes, or to a misspecification of the muscles. Two independent results indicated that Iro-C genes are required autonomously in the mesoderm to specify the LT fate. First, the normal development of LT muscles in Df(2L)5 embryos devoid of Iro-C gene expression at the ectoderm (Figure S2 and [44]). And second, the rescue of the muscle phenotype of Df(3L)iroDFM3 embryos by Ara supplied exclusively in the mesoderm (using myocyte enhancer factor 2 (mef2)-GAL4 as driver, Figure 3D).
All progenitors and founders segregate in Df(3L)iroEGP6 mutant embryos
We next examined whether the loss of LT muscles was due to either a failure in the segregation of muscle progenitors (absences and/or duplications) or to an early transformation of the fate of LT progenitors. To discern between these possibilities we combined the reporter line rP298, which expresses ß-galactosidase in all progenitors and founders [32], [45] with Df(3L)iroEGP6. We focussed on the previously well-established muscle lineages labelled by Slou/S59 [3], [17] and the LT1–4 lineages labelled by Kr [18]. With these markers in the lateral-ventral region of rP298 embryos we can identify the following founders (Figure 4A-4A″ and insets below). In the dorsalmost lateral mesoderm we find the sibling founders DT1 and DO3 (expressing slou) and the lateral longitudinal 1 (LL1) founder and its sibling (expressing Kr). Immediately below segregate the four LT founders (expressing Kr). And more ventrally appear the sibling ventral acute 1 (VA1) and VA2 founders (which express Kr and slou) and the VA3 founder and its sibling, the ventral adult precursor (that express slou). In Df(3L)iroEGP6 embryos we observed the same number of identifiable founders (Figure 4B and 4B′). There were however significant differences in terms of patterns of gene expression. Namely, the presumptive LT3–4 founders now expressed slou in addition to Kr (Figure 4B and 4B′ and insets below). This code of muscle identity gene expression is similar to that of VA1 and VA2 founders (Figure 5A), suggesting an early transformation of LT3–4 to VA1–2 muscles.


Ara and Caup implement LT muscle fate by repression of muscle identity genes in progenitors
The absence of all muscles with LT morphology in ara/caup mutant embryos prompted us to examine whether, in addition to the putative transformation of LT3–4 towards VA1–2, there was a similar change of fate for LT1–2. LT progenitors express Kr, caup, Con and lms, PLL1/LL1sib expresses Kr and vg, and PVA1/2 Kr, slou, Con and Poxm (Figure 5A and 5B). Using a combination of these markers we found in the lateral region of Df(3L)iroDFM3 embryos an ectopic muscle that expressed Kr+Vg, the code of LL1 (LL1*, Figure 5C) and an ectopic muscle VA2 (VA2* in Figure 5E–5G). This change of muscle identity could take place in founders or at the progenitor state. If this were the case, we anticipated that both muscles resulting from sibling founder myoblasts should be duplicated in Df(3L)iroDFM3 embryos. Indeed, using anti-Poxm, which labels VA1–3 ([14] and Figure 5B), and antibodies to Kr and Slou, which are maintained only in VA2 (Figure 5B and 5D), we identified two VA2 muscles (that co-express Poxm and Kr) and two Poxm-expressing VA1 muscles in late stage 14 Df(3L)iroDFM3 embryos (Figure 5F). The presence of the duplicated VA1 and VA2 muscles was more evident at stage 15 when Poxm was only weakly expressed in VA2 muscles (Figure 5G). We concluded that Ara and Caup were required to specify LT progenitors and that implementation of this fate implies the repression of specific muscle identity genes, such as slou in PLT3/4 and vg in PLT1/2. Moreover, it seemed that the only muscles affected by the lack of ara/caup were those in which these genes were already expressed in the corresponding promuscular clusters, since the fate of DT1 and SBM, visualised by the expression of slou, Con and lb, was apparently unaffected in Df(3L)iroDFM3 embryos (Figure 2F, Figure 5D and 5E, and Figure S1E–S1H).
Ras/MAPK cascade modulates the regulation of slou by Caup in Schneider-2 cells
Our data suggested that Ara/Caup might act as repressors of slou in the Drosophila mesoderm. Therefore we decided to investigate whether slou might be a direct target of Ara/Caup. An “in silico” search of a previously reported slou cis-regulatory region [46] identified two putative Iro binding sites (BS) at positions +129 (BS1) and −1642 (BS2), relative to the transcription start site, which match the consensus ACAN2–8TGT ([47] and Figure 6A). We cloned this regulatory region in a Luciferase reporter vector and measured Luciferase activity in Drosophila Schneider-2 (S2) cells transiently transfected with this construct and increasing amounts of HA-tagged Caup. Contrary to expectations, we found that addition of Caup-HA increased the basal Luciferase activity driven by the slou regulatory region in a dose dependent manner (blue bars in Figure 6B), indicating that Caup acts as a transcriptional activator of slou under these conditions. The reported regulation of the chicken Irx2 factor by MAPK (that switches it from repressor to activator) could explain this result [48]. Since Western Blot analysis of S2 lysates using an antibody against diphospho-extracellular-signal related kinase (dpErk) showed the MAPK pathway to be active in S2 cells (Figure 6C) and we have obtained experimental evidence showing the presence of phosphorylated Caup in S2 cells with constitutively active MAPK pathway (N.B, A.S.T and S.C, manuscript in preparation), we hypothesized that the activation effect of Caup in S2 cells could be due to the Ras/MAPK cascade turning Caup from transcriptional repressor into activator. Indeed, the inhibition of the Ras/MAPK pathway by the PD98059 MAP-erk kinase-1 (MEK1) inhibitor induced a Caup-dose dependent decrease in Luciferase activity driven by the slou regulatory sequences (Figure 6B, red bars). This result could not be attributed to a direct effect of the inhibitor over the slou promoter, since its addition did not modify the basal Luciferase activity of the construct (Figure 6B).

To test whether Caup-dependent transcriptional regulation relied on a direct interaction of Caup with the slou regulatory region we performed electrophoretic mobility shift assays (EMSA) with in vitro translated Caup and wild-type and mutated Caup-BS. These assays indicate efficient binding of Caup to BS1, which is abolished by BS1 mutation and deletion (Figure 6D). In contrast, Caup appears not to bind BS2 under these experimental conditions (not shown).
Next we examined the functional relevance of BS1 and BS2 in the Luciferase reporter assay. Deletion of BS2 had no major effect on Caup-dependent luciferase expression compared to the wild-type promoter (Figure 6E and 6F compare with Figure 6B). This result suggested that Caup does not bind to BS2 (as indicated by the EMSA data). Unexpectedly, deletion of BS1 resulted in a more efficient activation of luciferase expression than that driven by the wild type regulatory region (Figure 6E). This suggested that binding of Caup to BS1 somehow impaired transcription. Note that the activation of luciferase driven by the BS1 mutated regulatory region was still dependent on the MAPK pathway (Figure 6E and 6F). This suggests that such activation appears to depend on the binding of a MAPK-dependent phosphorylated protein, which we hypothesize might be Caup, to a so far unidentified binding site. Thus, the analysis in S2 cells confirmed the relevance of BS1, but not of BS2 on Caup-dependent regulation.
Additionally, we have analysed the evolutionary conservation of these putative Caup-BS among several Drosophila species (Figure S3). Notably, BS1 is located in a highly conserved region and its sequence is identical across the melanogaster group, whereas neither BS2 nor the adjacent sequences are conserved. These data further reinforce the relevance of BS1 for Caup-dependent slou regulation.
Our results are thus consistent with a direct effect of Caup on slou regulation. However, it cannot be ruled out the possibility of the existence in vivo of a transcription factor, acting downstream of ara/caup, that could repress slou through BS1 or through a still unidentified regulatory sequence of slou.
Caup integrates in vivo inputs from the Ras/MAPK cascade for its regulation of slou
To further examine in vivo the regulatory activity of Caup on slou (Figure 7B, 7C), we ectopically expressed caup or ara in VA1–3 using Con-GAL4 and checked whether they would repress slou in the VA2 muscle. This was indeed the case (Figure 7B, 7D, 7F-7F″ and not shown). Loss of slou expression caused by ectopic caup reproduced the morphological defects in VA2 previously described in slou mutants (Figure 7F-7F″ and [17]). To analyse whether the morphological effect of Caup on muscle VA2 development was only due to Caup-dependent repression of slou, we forced the expression of both genes using the Con-GAL4 driver. In this experimental condition Caup was unable to repress UAS-slou expression and the VA2 muscle and its morphology seemed unaffected (Figure 7F-7G″).

Once verified the repressor activity of Caup on slou during myogenesis, to analyse the regulatory potential of BS1 in vivo we generated transgenic flies harbouring the wild-type or the BS1 deleted version of the slou regulatory region. The wild-type regulatory region only partially reproduced the slou endogenous expression, as it drove lacZ expression in the CNS but not in the muscles (not shown and Figure S4). In contrast, the construct lacking BS1 behaved congruently with our S2 cells results, since it drove ectopic expression of lacZ in the lateral muscles (Figure S4). Curiously, up-regulation of lacZ was found in the 4 lateral muscles and not only in the ones that show slou expression in the absence of Ara/Caup (Figure 4B). Thus we interpret that this construct, while missing some of the regulatory sequences required for slou mesodermal expression, it contains those required for Caup mediated repression in the mesoderm. In addition, the absence of strict correlation between the phenotypes of deletion of BS1 and lack of Ara/Caup, might indicate the ability of other transcription factor(s) to regulate slou expression in LT1–2 through BS1.
To investigate whether the effect of the MAPK cascade on the transcriptional activity of Ara/Caup found in the S2 cell assay is also at work during myogenesis we examined whether there is a correlation between MAPK signalling and Caup transcriptional regulatory activity. We looked at the state of activation of this pathway in the LT promuscular cluster, where Ara/Caup repress slou, and found that it did not appreciably express dpErk (Figure 7H). Therefore, a repressor activity of Ara/Caup correlates in vivo with the absence of MAPK signalling. Next, we tested whether forced activation of the MAPK pathway in the mesoderm could interfere with the repressor activity of endogenous Caup in LT promuscular clusters. This was indeed the case, since activation of the MAPK pathway using twist-GAL4; 24B-GAL4 to drive the activated form of Ras85D (rasV12 [49]) allowed co-expression of caup and slou in this cluster (Figure 7I). Similarly, late co-expression of rasV12 and caup (Con-Gal4 driver) blocked the repression activity of Caup on slou (Figure 7D and 7E). Finally, to test whether MAPK signalling not only prevented Caup-dependent repression of slou but also converted Caup from repressor to activator, we looked at the expression of slou after early pan-mesodermal Caup expression (mef2-GAL4). As shown in Figure 7J, Ara was indeed able to ectopically activate slou in Drosophila epidermal growth factor receptor (DER)-dependent eve-expressing muscles.
Discussion
The study of myogenesis in Drosophila has increased the understanding of how the mechanisms that underlie the acquisition of specific properties by individual muscles are integrated within the myogenic terminal differentiation pathway. Thus, the current hypothesis proposes that distinct combinations of regulatory inputs leads to the activation of specific sets of muscle identity genes in progenitors that regulate the expression of a battery of downstream target genes responsible for executing the different developmental programmes (reviewed in [2], [10], [38]). However, the analysis of the specific role of individual muscle identity genes and of their hierarchical relationships is far from complete since the characterisation of direct targets for these transcriptional regulators is very scarce [36], [37].
Here, we report the identification of ara and caup, two members of the Iroquois complex, as novel type III muscle identity genes. We find that the homeodomain-containing Ara and Caup proteins are necessary for the specification of the LT fate. ara/caup appear to be bona fide muscle identity genes. Indeed, similarly to the identity genes Kr and slou [17], [18], absence of ara/caup does not interfere with the segregation of muscle progenitors or their terminal differentiation, but modifies the specific characteristics of LT1–4 muscles, which are transformed towards VA1, VA2, LL1 and LL1 sib fates. These transformations may be due in part to the up-regulation of slou and vg in the corresponding muscles. Thus, a recent report [50] shows that forced expression of vg in LT muscles induces changes in muscle attachments similar to the ones observed in LT1 in ara/caup mutant embryos. However, it should be stressed that although in ara/caup mutants LT muscles are lost in more than 95% of cases, they are not completely transformed into perfect duplicates of the newly acquired fates. For instance, while the specific LT marker lms is lost in 91% of cases, ectopic slou expression is detected in only 75% of cases. These partial transformations might be due to differences in the signalling inputs acting in the mesodermal region from where these muscles segregate (see below). Our unpublished data also showed that forced pan-mesodermal expression of ara/caup alter the fates of many muscles both in dorsal and in ventral regions without converting them into LT muscles (i.e., they do not ectopically express lms). Similarly, Kr and slou ectopic expression is not sufficient to implement a certain muscle fate [17], [18]. The failure to recreate a given muscle identity by adding just one of the relevant muscle identity proteins reveals the importance that cell context, that is, the specific combination of signalling inputs and gene regulators present in each cell, have in determining a specific muscle identity.
Our analysis of the myogenic requirement of ara/caup has revealed several features about how these genes act to implement LT fates. Thus, although they are expressed in six developing embryonic muscles, only four of them, LT1–4, are miss-specified in the absence of Ara/Caup. The remaining two, DT1 and SBM, seem to develop correctly, according to morphological as well as molecular criteria. It is worth noting that the requirement for ara/caup genes in these six muscles correlates with the onset of their expression. Thus, in the affected LT1–4 muscles Ara/Caup can be first detected at the earliest step of muscle lineages, that is in the promuscular clusters. In contrast, in the unaffected muscles ara/caup start to be expressed later, in the DT1/DO3 progenitor and the SBM founder. This suggests that in muscle lineages ara/caup have to be expressed very early to repress slou and vg to implement the LT fate. Several data support this interpretation. For instance, the observation that ara/caup are co-expressed with slou in DT1, whereas they repress slou in LT3–4, may be related to the fact that slou expression precedes that of ara/caup in the DT1 lineage. Should this be so, one would expect that ectopic expression of ara using the early driver mef2-GAL4, would repress slou in DT1, as it actually does (Figure S5), whereas this repression is not evident using the late driver Con-GAL4. Furthermore, the hypothesis of the relevance of the timing of muscle identity gene expression for muscle fate specification might also apply to the case of slou, where a similar correlation between the strength of the loss-of-function slou phenotypes in specific muscles and the onset of slou expression has also been found [17].
It should be stressed that the generation of the LT code depends not only on the early presence of Ara/Caup on the promuscular clusters but also on the absence (or strong reduction) of DER/Ras activity at that precise developmental stage and location (Figure 8). There is a dynamic regulation of MAPK signalling in the lateral mesoderm. Caup-expressing muscles develop from DER-independent clusters whereas the duplicated muscles observed in ara/caup mutants derive from progenitors that segregate very near the LT progenitors [3], but originate in DER-dependent promuscular clusters that are specified slightly later in development [4], [51]. Furthermore we have observed both by in vivo and in cell culture that low MAPK activity is required for Caup-dependent slou repression. Therefore, we interpret the role of Ara/Caup in the implementation of LT fate as follows (Figure 8). At mid stage 11 in the myogenic mesoderm, groups of mesodermal cells acquire myogenic competence as a result of interpreting a combinatorial signalling code that reflects their position along the main body axes, as well as the state of activation of different signalling pathways [4]. Accordingly, these clusters initiate the expression of lethal of scute and a unique code of muscle identity genes, as has been shown in great detail for eve expression in the dorsal mesoderm [34], [35]. In the case of the dorso-lateral mesoderm this code includes ara/caup and Kr and implements the LT fate. Since the level of activation of the Ras/MAPK cascade is low in these clusters, Ara/Caup will behave as transcriptional repressors, preventing the activation of slou or vg in LT1–2 and LT3–4 clusters, which would be otherwise activated in this location. Thus, Ara/Caup implement the LT fate by repressing the execution of the alternative fates (Kr+, Slou+, Con+, Poxm+ and Kr+, Vg+) that would give rise to duplicates of PVA1/VA2 and PLL1/LL1sib, respectively, and by allowing a different identity gene code (Kr+, Caup+, Con+, lms+) that generates the LT fate.

Slightly later the Ras/MAPK pathway becomes active at the dorsolateral region (Figure 8). This changes the combinatorial signalling code and coincides with a change in the muscle identity genes expressed by the promuscular clusters that segregate from this position, which now accumulate Kr but not Ara/Caup. Progenitors born from them will express either slou or vg and give rise to VA1–2 and LL1/LL1sib fates, all DER-dependent [51].
Our S2 cells experiments suggest a molecular mechanism by which the Ras/MAPK pathway modulates the transcriptional activity of Ara/Caup on slou. Thus, low MAPK activity and direct binding of Caup to BS1 site of the slou gene would favour strong repression of slou. BS1 could be embedded in a silencer regulatory element or its binding to Caup may block transcription of the downstream located luciferase gene. On the contrary, Caup-dependent activation of slou would be dependent on MAPK signalling. We hypothesize that MAPK–dependent Caup phosphorylation could modulate its interaction with different transcriptional co-factors or/and its binding site affinity.
Furthermore, our in vivo evidence indicates a repressor function of presumably non-phosphorylated Caup on slou since forced activation of the Ras pathway allows co-expression of slou and caup. On the other hand, the ectopic expression of slou induced by caup-over-expression is suggestive of a possible activator function of phosphorylated Caup.
The role of IRO proteins in cell fate specification is conserved in both vertebrates and invertebrates (reviewed in [52]). Here we have shown that the interplay between MAPK signalling and IRO activity found in vertebrate neuroepithelium [48] is also at work in Drosophila myogenesis. We have also identified a potential direct target of Ara/Caup, slou and propose vg as a candidate gene to be regulated by Ara/Caup. In both cases the genes subordinated to ara/caup encode transcription factors that might in turn regulate the expression of other genes, genes that must be repressed in LT muscles in order to acquire the LT fate. These results, therefore, provide insights into the way Ara/Caup control lateral muscle identity and on the role of signalling pathway inputs to modulate the activity of these transcription factors, with consequences in their downstream targets. It also highlights the importance that the specific combination of muscle identity genes, their hierarchical relationships and their temporal activation have in determining the identity of a given muscle cell, very alike to what is at work during the acquisition of neural fates [53].
Materials and Methods
Drosophila stocks
The following stocks were used: Df(3L)iroDFM3, ararF209 [28], Df(2L)5 [54], Df(3L)iroEGP6, Df(3L)iroEGP5, Df(3L)iroEGPΔ1 [42], rP298 [32], mef2-GAL4 [55], Con-GAL4 [56], twist-GAL4; 24B-GAL-4 (a gift from M. Baylies), UAS-ara, UAS-caup [28], UAS-caup-HA (N. Barrios, unpublished) and UAS-rasV12 [49]. Ectopic expression was generated by means of the GAL4/UAS system [57].
In situ hybridisation, immunohistochemistry, and microscopy
Whole-mount in situ hybridisation with digoxygenin-labelled RNA probes and immunocytochemistry were performed as described previously [58]. Stained embryos were embedded in Araldite and sectioned (3 µm) following standard procedures. The following primary antibodies were used at the indicated dilutions: rat anti-Caup (1∶50) [23], guinea pig anti-Kr (1∶500) [59], mouse anti-Lb (1∶1) [15], rabbit anti-Poxm (1∶10) [14], rat and rabbit S59 (that recognises Slou, 1∶50) [3], rabbit anti-Alien (1∶500) [60], mouse anti-Con (1∶10) [61], rabbit anti-Vg (1∶500) [62], rat-anti- Tropomyosin (MAC141; 1∶100; Babraham Tech), rabbit anti-Myosin (Myo; 1∶300) [63], rat anti-HA (1∶1000; Roche); rabbit anti-ß-Gal (1∶5000; Cappel) and mouse anti- dpErk (1∶50; Sigma). Images were obtained with confocal microscopes MicroRadiance (BioRad) and LSM510META (Zeiss) and analysed using the software Zeiss LSM Image or LaserSharp and Adobe Photoshop 7.0. In most cases the figures correspond to z-projections of series of confocal sections.
Cell culture and transfections
The 5′-upstream region of slou (from −1828 to +153 nt) was amplified via PCR and cloned in pGLHS43 vector, a modified version of the pGL2-basic vector (Luciferase reporter plasmid, Promega), obtained after substitution of the SV40 promoter by the Drosophila heat-shock 43 minimal promoter (a gift from A. Baonza). The putative Caup BS1 and BS2 were deleted using the “Quick Change” site-directed mutagenesis kit (Stratagene, SantaClara, CA). The sequences of the primers used to delete BS1 were 5′-GAGTTCTTAATCCAGCCGTGTTGTGTGCCTGTGGCAAGTCAATAG-3′ and its reverse complement and for BS2, 5′-CCATATACATATGTGTGCATGTATGCATAAGTGTGAGTGTGAGTGGG -3′ and its reverse complement. pAC5.1-Caup-HA plasmid was obtained after cloning caup ORF with an HA tag in the Drosophila expression vector pAC5.1 (Invitrogen). Drosophila S2 cells were cultured in Insect-Xpress medium (Lonza) supplemented with 7% fetal bovine serum and grown at 25°C. For Luciferase assays S2 cells were seeded at a density of 2×106 and co-transfected with 1 µg of the different firefly Luciferase reporter constructs DNA, 30 ng of control plasmid (expressing Renilla Luciferase driven by the promoter of Drosophila RpIII128, [64]) and either 0, 0.25, 0.5 or 1 µg of pAC5.1-Caupolican-HA plasmid per well using Nucleofector Technology (Lonza). Luciferase activity in the cell extracts was measured using Dual-Glo Luciferase assay system (Promega) following the manufacturer's protocol. Briefly, 20 µl extract was added to 100 µl F-luc assay reagent, mixed gently for 5 s and placed in a luminometer. After counting F-luc activity for 10 s, 100 µl stop-and-glo reagent was added to the tube, mixed gently for 5 s and placed in the luminometer for R-luc count. The R-luc activities were used as internal control to correct for the difference in transfection efficiency of different reporter plasmids. Therefore, F-Luc/R-Luc activities were used for data analysis. To investigate whether the MEK/ERK pathway was involved in transcriptional regulation driven by the slou promoter, S2 cells were treated or not with 50 µM PD-98059 (Sigma) for 2 hrs before Luciferase activity measurement. All data reported are means from three or four independent experiments, each performed in triplicates. Primary antibodies used in immunoblots were mouse anti-dpErk (1 µg/ml; Sigma), rat anti-HA (200 ng/ml; Roche) and mouse anti-βtubuline (1∶5000; Developmental Studies Hybridoma Bank).
Generation of slou reporter transgenic lines
The 5′-upstream region of slou used in S2 cells in the Luciferase reporter assays (both the wild type sequence and that missing the putative Caup BS1) were subcloned at the EcoRI site of the C4PLZ enhancer tester plasmid that contains a weak P-element promoter [65]. These lacZ reporter plasmids were introduced into y w1118 embryos by standard P-element transformation.
Electrophoretic mobility shift assay
Caup binding ability to the slou promoter region was analyzed by EMSA. Pairs of single-stranded, Cyc3 and unlabeled 40-mer oligonucleotides containing the wild-type putative Caup binding sites BS1, BS2 and their mutant or deleted versions were allowed to anneal to generate double-stranded probes. Sequences of primers are shown in Figure 6D for BS1 and in Dataset S1. Caup protein was synthesized in vitro by using the coupled transcription/translation rabbit reticulocyte lysate system (TNT Promega). The indicated amount of µl of TNT reaction mixture was incubated with 20 ng of labelled probe. Protein–DNA complexes were allowed to form at room temperature for 30 min in a total volume of 20 µl of binding buffer (50 mM HEPES, pH 7.5, 10 mM MgCl2, 10 mM KCl and 1 mM DTT). After incubation, free DNA and protein–DNA complexes were resolved by 6% non-denaturing polyacrylamide gel electrophoresis. Gel fluorescence was analyzed in a Typhoon Scanner (GE healthcare).
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. BateM 1993 The mesoderm and its derivatives; Martinez-AriasMBaA Cold Spring Harbor, New York CHS Laboratory Press 1013 1090
2. BayliesMKBateMRuiz GomezM 1998 Myogenesis: a view from Drosophila. Cell 93 921 927
3. CarmenaABateMJimenezF 1995 Lethal of scute, a proneural gene, participates in the specification of muscle progenitors during Drosophila embryogenesis. Genes Dev 9 2373 2383
4. CarmenaAGisselbrechtSHarrisonJJimenezFMichelsonAM 1998 Combinatorial signaling codes for the progressive determination of cell fates in the Drosophila embryonic mesoderm. Genes Dev 12 3910 3922
5. CarmenaAMurugasu-OeiBMenonDJimenezFChiaW 1998 Inscuteable and numb mediate asymmetric muscle progenitor cell divisions during Drosophila myogenesis. Genes Dev 12 304 315
6. Ruiz-GómezMBateM 1997 Segregation of myogenic lineages in Drosophila requires numb. Development 124 4857 4866
7. DuanHSkeathJBNguyenHT 2001 Drosophila Lame duck, a novel member of the Gli superfamily, acts as a key regulator of myogenesis by controlling fusion-competent myoblast development. Development 128 4489 4500
8. FurlongEEAndersenECNullBWhiteKPScottMP 2001 Patterns of gene expression during Drosophila mesoderm development. Science 293 1629 1633
9. Ruiz-GómezMCouttsNSusterMLLandgrafMBateM 2002 myoblasts incompetent encodes a zinc finger transcription factor required to specify fusion-competent myoblasts in Drosophila. Development 129 133 141
10. BayliesMKMichelsonAM 2001 Invertebrate myogenesis: looking back to the future of muscle development. Curr Opin Genet Dev 11 431 439
11. TixierVBatailleLJaglaK 2010 Diversification of muscle types: recent insights from Drosophila. Exp Cell Res 316 3019 3027
12. RushtonEDrysdaleRAbmayrSMMichelsonAMBateM 1995 Mutations in a novel gene, myoblast city, provide evidence in support of the founder cell hypothesis for Drosophila muscle development. Development 121 1979 1988
13. BourgouinCLundgrenSEThomasJB 1992 Apterous is a Drosophila LIM domain gene required for the development of a subset of embryonic muscles. Neuron 9 549 561
14. DuanHZhangCChenJSinkHFreiE 2007 A key role of Pox meso in somatic myogenesis of Drosophila. Development 134 3985 3997
15. JaglaTBellardFLutzYDretzenGBellardM 1998 ladybird determines cell fate decisions during diversification of Drosophila somatic muscles. Development 125 3699 3708
16. CrozatierMVincentA 1999 Requirement for the Drosophila COE transcription factor Collier in formation of an embryonic muscle: transcriptional response to notch signalling. Development 126 1495 1504
17. KnirrSAzpiazuNFraschM 1999 The role of the NK-homeobox gene slouch (S59) in somatic muscle patterning. Development 126 4525 4535
18. Ruiz-GómezMRomaniSHartmannCJackleHBateM 1997 Specific muscle identities are regulated by Kruppel during Drosophila embryogenesis. Development 124 3407 3414
19. BateMRushtonEFraschM 1993 A dual requirement for neurogenic genes in Drosophila myogenesis. Dev Suppl 149 161
20. NoseAMahajanVBGoodmanCS 1992 Connectin: a homophilic cell adhesion molecule expressed on a subset of muscles and the motoneurons that innervate them in Drosophila. Cell 70 553 567
21. Gomez-SkarmetaJLModolellJ 2002 Iroquois genes: genomic organization and function in vertebrate neural development. Curr Opin Genet Dev 12 403 408
22. CavodeassiFModolellJCampuzanoS 2000 The Iroquois homeobox genes function as dorsal selectors in the Drosophila head. Development 127 1921 1929
23. Diez del CorralRArocaPJLGm-SCavodeassiFModolellJ 1999 The Iroquois homeodomain proteins are required to specify body wall identity in Drosophila. Genes Dev 13 1754 1761
24. FeijooCGManzanaresMde la Calle-MustienesEGomez-SkarmetaJLAllendeML 2004 The Irx gene family in zebrafish: genomic structure, evolution and initial characterization of irx5b. Dev Genes Evol 214 277 284
25. CavodeassiFDiez Del CorralRCampuzanoSDominguezM 1999 Compartments and organising boundaries in the Drosophila eye: the role of the homeodomain Iroquois proteins. Development 126 4933 4942
26. GlavicAGomez-SkarmetaJLMayorR 2002 The homeoprotein Xiro1 is required for midbrain-hindbrain boundary formation. Development 129 1609 1621
27. Gomez-SkarmetaJde La Calle-MustienesEModolellJ 2001 The Wnt-activated Xiro1 gene encodes a repressor that is essential for neural development and downregulates Bmp4. Development 128 551 560
28. Gomez-SkarmetaJLDiez del CorralRde la Calle-MustienesEFerre-MarcoDModolellJ 1996 Araucan and caupolican, two members of the novel iroquois complex, encode homeoproteins that control proneural and vein-forming genes. Cell 85 95 105
29. LeynsLGomez-SkarmetaJLDambly-ChaudiereC 1996 iroquois: a prepattern gene that controls the formation of bristles on the thorax of Drosophila. Mech Dev 59 63 72
30. Rodriguez-SeguelEAlarconPGomez-SkarmetaJL 2009 The Xenopus Irx genes are essential for neural patterning and define the border between prethalamus and thalamus through mutual antagonism with the anterior repressors Fezf and Arx. Dev Biol 329 258 268
31. MichelsonAMAbmayrSMBateMAriasAMManiatisT 1990 Expression of a MyoD family member prefigures muscle pattern in Drosophila embryos. Genes Dev 4 2086 2097
32. NoseAIsshikiTTakeichiM 1998 Regional specification of muscle progenitors in Drosophila: the role of the msh homeobox gene. Development 125 215 223
33. DuboisLEnriquezJDaburonVCrozetFLebretonG 2007 Collier transcription in a single Drosophila muscle lineage: the combinatorial control of muscle identity. Development 134 4347 4355
34. KnirrSFraschM 2001 Molecular integration of inductive and mesoderm-intrinsic inputs governs even-skipped enhancer activity in a subset of pericardial and dorsal muscle progenitors. Dev Biol 238 13 26
35. PhilippakisAABusserBWGisselbrechtSSHeFSEstradaB 2006 Expression-guided in silico evaluation of candidate cis regulatory codes for Drosophila muscle founder cells. PLoS Comput Biol 2 e53 doi:10.1371/journal.pcbi.0020053
36. HartmannCLandgrafMBateMJackleH 1997 Kruppel target gene knockout participates in the proper innervation of a specific set of Drosophila larval muscles. Embo J 16 5299 5309
37. JunionGBatailleLJaglaTDa PonteJPTapinR 2007 Genome-wide view of cell fate specification: ladybird acts at multiple levels during diversification of muscle and heart precursors. Genes Dev 21 3163 3180
38. BatailleLDelonIDa PonteJPBrownNHJaglaK 2010 Downstream of identity genes: muscle-type-specific regulation of the fusion process. Dev Cell 19 317 328
39. CallejaMHerranzHEstellaCCasalJLawrenceP 2000 Generation of medial and lateral dorsal body domains by the pannier gene of Drosophila. Development 127 3971 3980
40. LetiziaABarrioRCampuzanoS 2007 Antagonistic and cooperative actions of the EGFR and Dpp pathways on the iroquois genes regulate Drosophila mesothorax specification and patterning. Development 134 1337 1346
41. McNeillHYangCHBrodskyMUngosJSimonMA 1997 mirror encodes a novel PBX-class homeoprotein that functions in the definition of the dorsal-ventral border in the Drosophila eye. Genes Dev 11 1073 1082
42. González-PérezE 2009 Función y regulación de las homeoproteínas del complejo génico Iroquois en los discos imaginales de ala y ojo-antena de Drosophila melanogaster Madrid Autónoma de Madrid 146
43. MullerDJaglaTBodartLMJahrlingNDodtHU 2010 Regulation and functions of the lms homeobox gene during development of embryonic lateral transverse muscles and direct flight muscles in Drosophila. PLoS ONE 5 e14323 doi:10.1371/journal.pone.0014323
44. HerranzH 2001 Estudio de la función del gen pannier en el desarrollo embrionario de Drosophila melanogaster Madrid Autónoma de Madrid 79
45. Ruiz-GómezMCouttsNPriceATaylorMVBateM 2000 Drosophila dumbfounded: a myoblast attractant essential for fusion. Cell 102 189 198
46. KimSJParkTLeeKK 1999 Identification of a novel cis-acting positive element responsible for the cell-specific expression of the NK-1 homeobox gene. Biochem Biophys Res Commun 257 538 544
47. BilioniACraigGHillCMcNeillH 2005 Iroquois transcription factors recognize a unique motif to mediate transcriptional repression in vivo. Proc Natl Acad Sci U S A 102 14671 14676
48. MatsumotoKNishiharaSKamimuraMShiraishiTOtoguroT 2004 The prepattern transcription factor Irx2, a target of the FGF8/MAP kinase cascade, is involved in cerebellum formation. Nat Neurosci 7 605 612
49. LeeTFeigLMontellDJ 1996 Two distinct roles for Ras in a developmentally regulated cell migration. Development 122 409 418
50. DengHBellJBSimmondsAJ 2010 Vestigial is required during late-stage muscle differentiation in Drosophila melanogaster embryos. Mol Biol Cell 21 3304 3316
51. BuffECarmenaAGisselbrechtSJimenezFMichelsonAM 1998 Signalling by the Drosophila epidermal growth factor receptor is required for the specification and diversification of embryonic muscle progenitors. Development 125 2075 2086
52. CavodeassiFModolellJGomez-SkarmetaJL 2001 The Iroquois family of genes: from body building to neural patterning. Development 128 2847 2855
53. BaumgardtMMiguel-AliagaIKarlssonDEkmanHThorS 2007 Specification of neuronal identities by feedforward combinatorial coding. PLoS Biol 5 e37 doi:10.1371/journal.pbio.0050037
54. de CelisJFBarrioRKafatosFC 1996 A gene complex acting downstream of dpp in Drosophila wing morphogenesis. Nature 381 421 424
55. RanganayakuluGElliottDAHarveyRPOlsonEN 1998 Divergent roles for NK-2 class homeobox genes in cardiogenesis in flies and mice. Development 125 3037 3048
56. CallejaMMorenoEPelazSMorataG 1996 Visualization of gene expression in living adult Drosophila. Science 274 252 255
57. BrandAHPerrimonN 1993 Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118 401 415
58. MartinBSRuiz-GomezMLandgrafMBateM 2001 A distinct set of founders and fusion-competent myoblasts make visceral muscles in the Drosophila embryo. Development 128 3331 3338
59. KosmanDSmallSReinitzJ 1998 Rapid preparation of a panel of polyclonal antibodies to Drosophila segmentation proteins. Dev Genes Evol 208 290 294
60. GoubeaudAKnirrSRenkawitz-PohlRPaululatA 1996 The Drosophila gene alien is expressed in the muscle attachment sites during embryogenesis and encodes a protein highly conserved between plants, Drosophila and vertebrates. Mech Dev 57 59 68
61. MeadowsLAGellDBroadieKGouldAPWhiteRA 1994 The cell adhesion molecule, connectin, and the development of the Drosophila neuromuscular system. J Cell Sci 107 Pt 1 321 328
62. WilliamsJABellJBCarrollSB 1991 Control of Drosophila wing and haltere development by the nuclear vestigial gene product. Genes Dev 5 2481 2495
63. KiehartDPFeghaliR 1986 Cytoplasmic myosin from Drosophila melanogaster. J Cell Biol 103 1517 1525
64. GilchristDANechaevSLeeCGhoshSKCollinsJB 2008 NELF-mediated stalling of Pol II can enhance gene expression by blocking promoter-proximal nucleosome assembly. Genes Dev 22 1921 1933
65. WhartonKAJrCrewsST 1993 CNS midline enhancers of the Drosophila slit and Toll genes. Mech Dev 40 141 154
Štítky
Genetika Reprodukčná medicínaČlánok vyšiel v časopise
PLOS Genetics
2011 Číslo 7
- Je „freeze-all“ pro všechny? Odborníci na fertilitu diskutovali na virtuálním summitu
- Gynekologové a odborníci na reprodukční medicínu se sejdou na prvním virtuálním summitu
Najčítanejšie v tomto čísle
- Genome-Wide Association Study Identifies Novel Restless Legs Syndrome Susceptibility Loci on 2p14 and 16q12.1
- Loss of the BMP Antagonist, SMOC-1, Causes Ophthalmo-Acromelic (Waardenburg Anophthalmia) Syndrome in Humans and Mice
- Gene-Based Tests of Association
- Genome-Wide Association Study Identifies as a Susceptibility Gene for Pediatric Asthma in Asian Populations