
Základy kognitivní, afektivní a sociální neurovědy
VI. Volní rozhodování
Basics of cognitive, affective and social neuroscience. VI. Free will
Results of neurobiological experiments with animals and humans have become part of the discourse about free will in the new millenium. There are two neuronal circuits of volition in human brain:
– ventral (whose nodes are basal ganglia – prefrontal cortex – presupplemental motor area – supplementary motor area – primary motor cortex), and
– dorsal (whose nodes are sensory cortices – parietal cortex – lateral promotor cortex – primary motor cortex).
The anterior insular cortex is a newly discovered node. Phases of volition are
– the early whether decision,
– what decisions (goal selection and movement selection) , and
– the late whether decision (final predictive check and veto);
– when decision.
Damage of those nodes / phases is recognised in a number of common ailments.
Unconscious mechanisms widely influence free will. Free will is an adaptive biological trait. The scientific concept of free will is a quantitative not qualitative concept. The right question is probably “how much free will do have we?”, not “do we have free will or not?”
Key words:
free will, neuronal circuits, unconscious mechanisms, adaptive trait.
Authors:
F. Koukolík
Authors‘ workplace:
Primář: MUDr. František Koukolík, DrSc.
; Oddělení patologie a molekulární medicíny
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
Published in:
Prakt. Lék. 2011; 91(6): 315-320
Category:
Editorial
Overview
Novou součástí tisícileté diskuse o svobodné vůli jsou výsledky neurobiologických experimentů se zvířaty i lidmi. Neuronální obvody volního rozhodování jsou v lidském mozku dva:
– ventrální, (jehož uzly jsou bazální ganglia – prefrontální kůra – presuplementární kůra – doplňková motorická oblast – primární motorická kůra), a
– dorzální, (jehož uzly jsou senzorické korové oblasti – parietální kůra – laterální premotorická kůra – primární motorická kůra).
Nově objeveným uzlem je kůra přední insuly. Fáze rozhodovacího procesu jsou
– rané rozhodnutí zda?,
– rozhodnutí co? (volba cíle spjatá s rozhodnutím o pohybu), a
– pozdní kontrola včetně případného veta;
– rozhodnutí kdy.
Poškození neuronální uzlů, respektive fází rozhodovacího procesu je známo u řady běžných onemocnění.
Svobodnou vůli rozsáhle ovlivňují nevědomé mechanismy. Svobodná vůle je adaptivní biologický znak. Vědecký pojem svobodné vůle je kvantitativní, nikoli kvalitativní pojem. Správně položená otázka pravděpodobně zní: „Kolik svobodné vůle máme?“, nikoli otázka „Máme, nebo nemáme svobodnou vůli?“.
Klíčová slova:
svobodná vůle, neuronální obvody, nevědomé mechanismy, adaptivní znak.
Úvod
Volní rozhodování a s ním spjatá otázka svobodné vůle je předmětem debaty filozofů a teologů od antiky, koncem 19. století se k diskusi přidala psychologie, v šedesátých letech 20. století kognitivní věda. Proslulým experimentem Benjamina Libeta vstoupila r. 1983 do problematiky neurověda. Pro filozofy, lékaře a patrně i neurovědce nečekaně a rozsáhle diskusi obohatily interpretace desítky let shromažďovaných výsledků experimentální práce s bezobratlými živočichy (19, 6).
Následující stať se zaměřuje na stručný a výběrový přehled současných výsledků neurovědy a biologie. Filozofickou problematiku, která je námětem bezpočetných knih a časopiseckých publikací (14), stať ponechává stranou. V tabulce 1 (str. 319) jsou pojmy často užívané filozofy v debatách o svobodné vůli a volním rozhodování, některé z nich se objevují v textu.

Současné poznatky neurovědy a biologie
V průběhu Libetových experimentů (24) pozorovali účastníci pokusů pohyb světelné skvrny nebo ručičky na hodinovém číselníku. Elektrickou aktivitu jejich mozku přitom snímal elektroencefalograf. Jakmile se sami spontánně rozhodli, pohnuli pravým zápěstím. Hodiny se v náhodném intervalu poté zastavily. Účastnici pokusu sdělili, ve které poloze byla ručička, respektive skvrna v okamžiku, kdy prvně „pocítili nutkání“, neboli uvědomili si záměr pohnout rukou, což se považuje za okamžik vědomého volního rozhodnutí., „svobodné vůle“, říkající: „Pohnu pravým zápěstím“. V průměru to bylo 206 ms před počátkem svalové aktivity.
Okamžik, kdy pokusné osoby prožily vědomí záměru (volní akce) označil Libet písmenem W (will). Elektrická aktivita mozku (Bereitschaftspotenzial, readiness-potential, přípravný potenciál, BP) odpovídající pohybu, se však objevovala přibližně 1 s před počátkem pohybu. Účastníkův mozek tedy připravoval akci dřív, než si účastník začal být vědom, že jeho záměrem je pohnout rukou. Okamžik, kdy subjekt pohnul zápěstím, označil Libet písmenem M (movement).
Interpretace pokusu byla kritizována filozofy, například Danielem Dennettem (9), podle jehož názoru je výsledek pokusu dán přesunem pozornosti od záměru na hodinový číselník, což je příčinou časového posunu mezi prožívanou zkušeností volního rozhodnutí a vnímanou polohou ručičky na číselníku. Ovlivnění výsledku experimentu pozorností následné experimenty potvrdily, nicméně základní zjištění, že činnost mozku předchází vědomému záměru, zůstalo v platnosti.
Haggard a Eimer (16) experiment modifikovali: pokusné osoby se kromě záměru pohybu rozhodovaly, zda užijí levou, nebo pravou ruku. Zkušenost záměru („chci pohnout rukou“) v tomto případě lépe korelovala se stranovým BP, oznamujícím, která hemisféra řídí pohyb. Z toho plyne, že vědomé volní rozhodnutí následuje až po přecházející elektrické aktivitě mozku, stejně jako v Libetově experimentu.
Banks a Isham (2) modifikovali Libetův experiment dvěma způsoby.
V první podobě poskytli pokusným osobám falešnou zvukovou zpětnou vazbu 5–60 ms po skutečném započetí motorické akce. Pokusné osoby úměrně tomu posunuly v čase dopředu dobu, kdy se „volně rozhodly“. Posun pocitu jejich „volního rozhodnutí“ odpovídal době odstupu falešné zpětné vazby.
V druhé podobě sledovaly pokusné osoby svou vlastní ruku na videozáznamu, jenž buď neměl, nebo měl zpoždění 120 ms; v tomto případě tedy opět poskytoval falešnou zpětnou vazbu o čase, v němž začínala motorická akce. Pokusné osoby stejně jako v předchozím experimentu při falešné zpětné vazbě posunuly v čase dopředu pocit vlastního volního rozhodnutí, a to o 44 ms.
Jinak řečeno: na okamžik v němž se k nějakému činu rozhodujeme, usuzujeme ze zdánlivého okamžiku odpovědi, v čase ho přesně nevnímáme. Okamžik volního rozhodnutí W je tedy introspektivní iluze, což je experimentálně dobře doložený jev.
Například Kühn a Brass (23) ukázali, jak některé pokusné osoby mají za to, že záměrně spustily volní akci, přičemž z reakčního času plyne, že ji ve skutečnosti nedokázaly zastavit.
Experiment Soonovy skupiny (30) užil při modifikovaném Libetově experimentu funkční magnetickou rezonanci. Pokusné osoby sledovaly proměny písmen, na které se dívaly. Písmena, například k, t, d, q, se měnila každých 500 ms. V některém z okamžiků se při pozorování písmene pokusné osoby samy rozhodly, že levým, nebo pravým ukazováčkem stisknou tlačítko („svobodná odpověď“). Následně jim byla předvedena časová mapa jejich odpovědí spolu s instrukcí oznámit stiskem tlačítka okamžik, v němž se ke „svobodné odpovědi“ rozhodli.
Experiment dokázal, že pocitu rozhodnutí předcházela aktivita prefrontální a parietální kůry až 10 s před tím, než se pocit svobodného rozhodnutí objevil ve vědomí.
Neuronální obvody volní akce
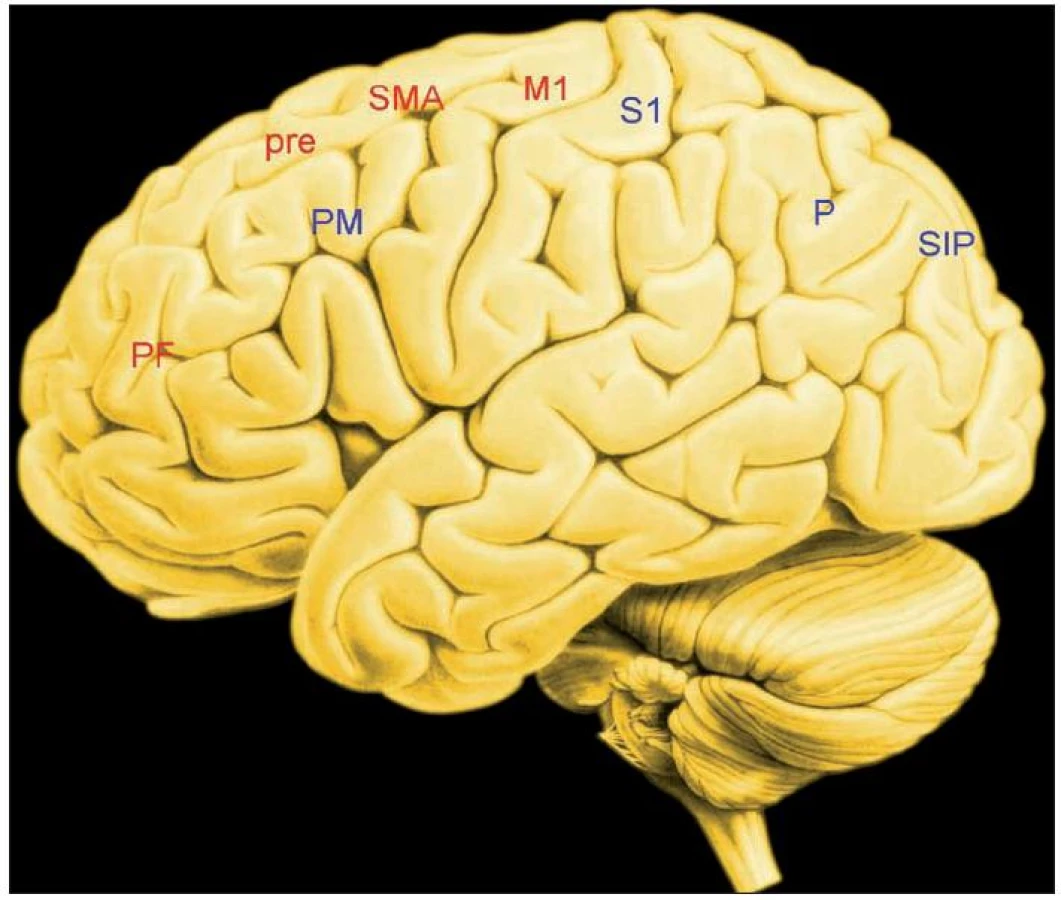
„Vykonavatelem“ volních akcí je primární motorická kůra M1. Dostává dva rozsáhlé vstupy:
1. Zdrojem prvního je prefrontální kůra (a její propojení s bazálními ganglii), která promítá do presuplementární kůry (pre-SMA), a to spolu se suplementární motorickou kůrou do M1. Pre-SMA se v jednoduchém experimentu rozlišujícím mezi pohyby ukazováčku na základě povelu a jeho pohyby na základě vlastního rozhodnutí aktivuje víc, jde-li o vlastní rozhodnutí (přehled 21). Pre-SMA je kromě toho součástí rozsáhlé kognitivní a motorické sítě, jejímiž uzly jsou premotorická, frontální polární a cingulární kůra. Pre-SMA je zdrojem rané fáze BP (29). Raná fáze BP je na začátku kaskády oslovující vlastní doplňkovou motorickou oblast (SMA) a následně M1.
Otázkou je, co spustí ranou fázi BP v pre-SMA. S vysokou pravděpodobností je to aktivita bazálních ganglií a jejich projekce do pre-SMA (1). Kromě klinické zkušenosti s omezením pohybů u pacientů s Parkinsonovou nemocí pro to svědčí signály předpovídající spuštění volní akce objevující se v elektrodách implantovaných do bazálních ganglií 2 před započetím pohybu.
Jinak řečeno: spuštění volní akce je projevem činnosti „obvodu“, „smyčky“ bazální ganglia – prefrontální kůra – pre-SMA – M1, nikoli hierarchizovaného systému, na jehož vrcholu je blíže nespecifikovaná a na ostatním nezávislá „vůle“.
Popsanou projekci lze popsat jako ventrální.
2. Zdrojem druhého vstupu jsou senzorické korové oblasti, a to prostřednictvím temenních laloků promítající do laterální oblasti premotorické kůry, ta na oplátku promítá do M1. Příkladem činnosti této sítě je cílený úchop pozorovaného objektu. Parietální oblast kolem sulcus intraparietalis se pravděpodobně podílí na volbě alternativ, tedy na volním rozhodování.
Tuto projekci je možné popsat jako dorzální (obr. 1.)
Volní akce jako rozhodování
Experimenty Libetova typu mluví o bezprostředních jednoduchých rozhodnutích. Subjekt přitom ví, že má učinit nějakou akci a je na jeho rozhodnutí, kdy ji učiní. Do rámce problematiky volní akce a svobodné volby patří však i dlouhodobější akce, které je možné rozdělit do několika fází, respektive navazujících rozhodovacích procesů:
- rané rozhodnutí „zda?“,
- rozhodnutí „co?“, neboli volbu cíle, spjaté s volbou příslušného pohybu,
- pozdní rozhodnutí „zda?“, neboli závěrečný odhad výsledku akce, včetně případného rozhodnutí o vetování akce,
- rozhodnutí „kdy?“ (17).
Rané rozhodování „zda?“
může být spuštěno tím, že
- a) rutinní zpracovávání informací na volbu odpovědi nestačí, například při rozhodování o alternativách plynoucích z nejednoznačných podnětů,
- b) objeví se nový důvod pro akci, třeba pocit žízně,
- c) jde o zkoumání, subjekt testuje další možnosti chování.
Rozhodnutí „co?“
neboli volba, cíle obvykle rozhoduje mezi alternativami. Poškození této části rozhodovacího procesu se objevuje (přehled 18):
- u lidí s poruchou řídících funkcí (dysexekutivní syndrom) v důsledku poškození zejména zevní prefrontální kůry, kromě jiného vázne uspořádávání akcí v časovém pořadí;
- choreu charakterizují volně nekontrolovatelné pohyby v raných fázích onemocnění, která jsou příčinou syndromu, své pohyby nemocní za mimovolní často nepovažují;
- pacienti s tiky například při Tourettově syndromu, často nejsou schopni rozlišit, zda jejich pohyb byl, nebo nebyl volně kontrolovaný;
- akinetický mutismus je podmíněn oboustrannou lézí vnitřní frontální kůry postihující SMA, pre-SMA a motorické oblasti g. cinguli;
- syndrom odcizené, respektive anarchické ruky podmíněný jednostrannou lézí vnitřní frontální kůry, obvykle SMA, pre-SMA: syndrom je mnohotvárný, pacienti mají například pocit, že jim končetina nepatří, ruka přitom vykonává anarchické pohyby, které se pletou do činnosti volně kontrolované ruky. Pacienti situaci někdy řeší tím, že v bdělém stavu na postižené končetině sedí, na noc ji uvazují.
- při disociativních syndromech;
- při psychogenním tremoru;
- v rámci syndromu fantomové končetiny;
- v rámci schizofrenního bludu: pacienti mají pocit, že jimi pohybuje někdo cizí nebo něco cizího.
Čelní laloky a zejména pre-SMA jsou tedy oblastí, která spojuje záměr s akcí. Volní rozhodování mezi alternativami a „přepínání“ mezi jednotlivými alternativami aktivuje vymezené oblasti kaudální a rostrální pre-SMA (26).
Součástí rozhodnutí „co?“ jsou rozhodnutí o pohybu, jimiž se cíl dosahuje, tato rozhodnutí aktivují temenní a premotorické oblasti. Například rozhodnutí o užití levé nebo pravé ruky aktivují polární frontální kůru, přes-SMA a rostrální cingulární kůru. Temenní a premotorické korové oblasti jsou aktivovány rozhodováním o specifické prostorové akci.
Pozdní rozhodnutí „zda?“,
neboli závěrečný odhad výsledku akce, včetně případného rozhodnutí o vetování akce: v době počátečního rozhodnutí často nelze předvídat, jak akce dopadne. Vyústění může být nejisté, případně riskantní, například proto, že se v průběhu akce změnil kontext. Akci je nutné korigovat, případně zastavit („vetovat“).
Brass a Haggard (3) instruovali pokusné osoby, aby jednak volní akci spustily, jednak aby volní akci na poslední chvíli zastavily. V obou případech pokusné osoby oznamovaly okamžik, v němž se rozhodly. Kontrolou byl zvuk, pokusné osoby určovaly, kdy zazněl. Inhibice volní akce je korelována aktivitou přední mediální prefrontální kůry rostrálně („vepředu“) od pre-SMA a přední insulární kůry.
Význam činnosti přední insulární kůry pro volní rozhodování začíná být doceňován až v poslední době. Vysvětlují ji dvě hypotézy:
- první má za to, že se přední insulární kůra podílí na tvorbě záměrů tím, že coby uzel sítě, která zpracovává informace z vnitřního prostředí, informuje o jeho stavu.
- druhá, podle autorů přehledné studie pravděpodobnější hypotéza, uvažuje o opaku: přední insulární kůra vyhodnocuje výsledky rozhodnutí o záměrné akci, rozhodnutí přitom formovala odlišná síť. Přední insulární kůra integruje interoceptivní signály, které jsou pro vyhodnocení důsledků akce klíčové (4).
Rozhodnutí „kdy?“
rozdíl mezi jednoduchou motorickou akcí, kterou pokusná osoba dělá na základě zevního signálu a akcí, kterou dělá na základě vlastního rozhodnutí (tu je možné chápat jako rozhodnutí „kdy?“) doprovází rozdíly aktivity pre-SMA a dorzolaterální prefrontální kůry. Časování akce je možné předpovědět z přípravné činnosti pre-SMA (30)
V podmínkách běžného života, nikoli jednoduchého laboratorního experimentu, je však časování akce podstatně složitější. Načasování volní akce závisí jak na prostředí, tak na motivacích.
Volní rozhodování a sebeuvědomování
Stručně a výběrově popsané experimenty dokazují, že elektrická aktivita mozku, která je podkladem volního pohybu, předchází okamžik, v němž si volní rozhodnutí k pohybu uvědomujeme. Přitom máme jednoznačný subjektivní pocit, že jsme strůjci své akce. „Vůle“ je podle tohoto pocitu jejím autorem. Podle této představy (svobodná vůle – spuštění mozkových motorických mechanismů – pohyb) je však vůle deus in machina, „homunculus“, „vrchol pyramidy“, výsledkem je nějaký druh dualistického pojetí vztahu mozek – vědomí (sebeuvědomování).
Přestala proto svobodná vůle (neurovědecky) existovat?
Jestliže chápeme svobodnou vůli jako druh percepce, nikoli autonomní rozhodovací mechanismus, pak nikoli (18). Popsané experimenty pro tuto možnost svědčí. Jak vědomý záměr (a rozhodnutí), tak motorická aktivita jsou projevem paralelní, byť v čase vzájemně posunuté aktivity mozku. Sebeuvědomování (mind) usuzuje na kauzální vztah záměru a akce ve snaze vysvětlit jejich korelaci. Jak záměr, tak akce mají společný jmenovatel: neuronální přípravu k akci (31, 32).
„Lidé prožívají svobodnou vůli, jestliže interpretují své vlastní myšlení jako příčinu své akce“ (31) je výrok dvou psychologů, Wegnera a Wheatleyové z r. 1999, který navazuje na úvahu Davida Humea, skotského filozofa z r. 1739, který říká, že svobodná vůle není „nic jiného než niterný dojem, který pociťujeme a jehož si jsme vědomi, jakmile vědomě podnítíme jakýkoli nový pohyb svého těla nebo nový vjem své mysli“ (20).
Podobný názor dokazuje, že vědomý záměr je dedukce vložená zpětně do proudu vědomí coby hypotetická příčina pohybu našich těl (10). Na tuto možnost ukazuje často bludné prožívání vztahu záměru a akce v průběhu neléčené schizofrenie.
„Nevědomá vůle“
Jakmile jsou dány cíle, považujeme svá rozhodnutí obvykle za vědomá. Experimenty ukazují, v jakém rozsahu jsou naše „svobodná volní rozhodnutí“ ovlivňována nevědomými mechanismy (přehled 7).
Jeden z velkého počtu příkladů: jak známo, pro většinu lidí jsou mohutnou motivací peníze. Pessiglione et al. (27) lidem předváděli obrázky mincí, a to jedné pence, a obrázek jejího stonásobku, jedné libry.
Instrukce zněla:
„Jakmile se podíváte na obrázek mince, stiskněte rukojeť. Čím silněji stisknete, tím bude váš finanční podíl větší.“
Promítání obrázku se přitom opakovalo mnohokrát, a to náhodně. Sílu své odpovědi lidé spatřili na stupnici podobné teploměru. Na posledním promítnutém obrázku pak zjistili, kolik si vydělali.
Obrázky byly promítány dvojím způsobem. Jednak tak, že lidé měli dost času rozlišit, kterou minci vidí, jednak velmi krátkou dobu, a to 17, nebo 50, nebo 100 milisekund, přičemž projekci obrázku na 100 milisekund předcházela optická „maska“ složená z obrázků obou mincí. Maska promítání i následovala, a to po dobu 100 milisekund zkrácenou o dobu vlastní projekce mincí. Tak krátké promítání sice mozek zaznamená, ale do vědomí se nedostane, podnět je podprahový. Lidé podprahový podnět nerozlišují, na otázku, co spatřili, odpovídali, že viděli masku.
V průběhu experimentu byla pokusným osobám sledována činnost mozku a elektrická kožní vodivost. Pohled na mince elektrickou kožní vodivost změnil. To znamená, že peníze jsou podnětem, který pro tyto lidi má emoční význam. Jestliže se lidé dívali a věděli, na co se dívají, byla síla stisku při librové minci větší než při pohledu na penci. Jestliže jejich mozkem prošel obrázek librové mince a pence nevědomě, opět byla síla jejich stisku větší při librové minci, než při nevědomém zpracovávání obrázku jedné pence, byť byl stisk slabší, než při vědomém pozorování.
Funkční magnetická rezonance našla jen jedinou mozkovou oblast, kterou obrázek libry aktivoval víc než pence – a tou bylo ventrální palidum, spodní oblast bazálních ganglií mozku a jeho okolí. Ventrální palidum se stejně jako nucleus accumbens (ten je v sousedství) podílí na tvorbě pocitu slasti. Je to zřejmě tahle oblast mozku, která vypočte očekávanou slast a přeloží ji do síly stisku. Rozdíl činnosti mozku v průběhu vědomé a nevědomé motivace vědci byl jen kvantitativní: při nevědomé motivaci byla aktivita, a tedy i síla stisku slabší.
Svobodná vůle jako biologická vlastnost
„V biologii nemá smysl nic, není-li to ve světle evoluce“, napsal dva roky před smrtí Theodosius Dobzhansky (1900–1975; 8). Někteří soudobí biologové mají za to, že svobodná vůle je „normální biologická vlastnost, nejde o dar ani tajemství“ (12, 13, 6, 19), což bude pravděpodobně překvapovat filozofy.
Modely, které se v této souvislosti studují, jsou únikové reakce. Příkladem předpověditelné odpovědi je C-startovní odpověď ryb. Ryby, které rozliší náhle zvýšený tlak z jedné strany těla, ohnou tělo do tvaru písmene C odvráceného od podnětu a snaží se uniknout. Vodnářka tykadlová (Erpeton tentaculatus), vodní had, vyvolává tuto předpověditelnou odpověď rybek pohybem kaudální části těla, takže rybky obratem vyrazí přímo do její tlamy.
Předpověditelnou odpověď much dobře maskovaných v povrchovém krytu místa kde pobíhají, podobně provokuje i lednáček bělokřídlý (Myioborus pictus). Mouchy vyskočí, pták je snadno rozliší proti nebi, takže se stanou kořistí.
Deterministické, předpověditelné chování však nemůže být evolučně stabilní. Selekční tlak favorizuje nepředpověditelnost. Živočich, který se chová předpověditelným způsobem, má nižší naději na přežití než živočich, který se chová nepředpověditelně. Variabilita chování je tudíž adaptivní vlastnost. Obecně viděno živočiši musí uvést do rovnováhy efektivitu svého chování s natolik dostatečnou mírou proměnlivosti, že sníží předpověditelnost. Efektivita odpovědí je kontrolována prostředím. Niterně kontrolovaná variabilita může snižovat efektivitu, zato sníží míru předpověditelnosti (přehled 6).
Jestliže ve dvou uvedených příkladech je chování živočichů předpověditelné, příkladem malé předpověditelnosti je chování švábů. V zadní spodní části těla mají cerci, dva vláskovité útvary, na každé straně jeden, což jsou čidla detekující pohyby vzduchu. Jakmile je čidla rozliší, spustí únikovou reakci, která šváby vede od místa, kde pohyb rozlišili. Úhel úniku ve vztahu ke směru vzdušného pohybu je však vysoce proměnlivý, takže směr úniku je pro predátora určitelný obtížně (11).
Filozofický spor o svobodnou vůli bývá dichotomický – strany sporu jsou buď zastánci determinismu, nebo indeterminismu. Patrně jde o falešnou dichotomii. Pozorování i experimenty ovlivňující chování zvířat ukazují, že jejich chování není zcela náhodné a přitom je špatně předvídatelné. Mozky zvířat a lidí lze chápat jako nestabilní nelineární systémy vysoce citlivé na nepatrné odchylky vstupních podmínek. Následné zesilování odchylek (amplifikace) je příčinou případně exponenciálně odlišného výsledku činnosti stejných systémů, jejichž vstupní podmínky byly na počátku téměř stejné (6). Příkladem takové amplifikace je chování sloupců senzorické kůry (25).
Jestliže chápeme proměnlivost a malou předpověditelnost chování jako adaptivní znak, pak je nutné odlišovat odpovědi od akcí. Akce jsou druhy chování, u nichž nelze zjistit vyvolávací podnět, nebo je latence a mohutnost odpovědi natolik proměnlivá, že je pojem odpověď na podnět bezcenná. Experimenty s octomilkami ukazují, že jejich chování je daleko variabilnější, než by odpovídalo variabilitě činnosti neuronů, které zprostředkovávají reakci podnět – odpověď (6). Byť se v prostředí neděje nic, co by je nutilo měnit chování, uchovávají si jeho variabilitu. Každá z proměn je spontánní akce, z nitra systému, není odpovědí na spouštějící zevní podnět.
Příkladem může být izolovaný nervový systém pijavky, který na základě neměnného elektrického podnětu někdy volí motorický program, odpovídající plavání, jindy lezení (15, 22). V tomto případě signály, které by mohly uniknout experimentátorově pozornosti, neexistují, proměny chování vznikají v systému samotném.
Chápat zdánlivě jednoduché živočichy jako biologické roboty by bylo zcela chybné, aniž je k tomu nutná mystika. Octomilky, stejně jako jiní živočiši a lidé, neznají řešení všech problémů předem. Nízká předpověditelnost jejich chování zvyšuje pravděpodobnost úniku před predátory a úspěchu v soutěži s příslušníky vlastního druhu, kromě toho hledají různá řešení nepředvídaných a nepředvídatelných problémů. Čím víc je „pasivní“ odpověď nahrazována akcí, tím větší pravděpodobnost evolučního úspěchu: přežití a potomstva.
Metafyzický pojem svobodné vůle / volního rozhodování rozlišuje krajnosti:
- naprostá náhoda / naprostá nutnost,
- čistý indeterminismus / tvrdý determinismus.
Pravděpodobnost (stochasticita) však není strašidlo, je součástí skutečnosti.
Vědecký pojem svobodné vůle je kvantitativní, nikoli kvalitativní pojem. Otázka není správně položená, ptáme-li se:
„Máme, nebo nemáme svobodnou vůli?“, ale: „Kolik svobodné vůle jsme měli, máme nebo budeme mít?“.
Dostává se tedy mezi krajnosti metafyzického pojetí (5, 6, 19).
John Searle, filozof, popsal svobodnou vůli jako přesvědčení, že „jsme se často mohli chovat jinak, než jsme se ve skutečnosti chovali“ (28). Pijavky i drosofily se dokáží chovat ve stejném prostředí odlišně. Tento druh svobody je nutnou, nikoli však postačující složkou svobodné vůle. Variabilita, respektive stochasticita chování je sama o sobě jen první složkou. Následuje rozhodnutí o tom, kterou z nabízených možností živočich nebo člověk volí. První krok je „svobodný“, druhý krok je „volní“, což odpovídá dvousložkovému modelu svobodné vůle navrženému jako prvním Williamem Jamesem a prosazovaném později Poincarém, Comptonem, Popperem, Dennetttem, Kanem, Fisherem, Melem, Kosslynem, Doylem, Kochem a Heisenbergem (přehled 6, 13).
MUDr. František Koukolík, DrSc.
Fakultní Thomayerova nemocnice s poliklinikou
Oddělení
patologie a molekulární
medicíny
Národní
referenční laboratoř prionových
chorob
Vídeňská
800
140
59 Praha 4 Krč
E-mail:
frantisek.koukolik@ftn.cz
Sources
1. Akkal, D., Dum, R.P., Strick, P.L. Supplementary motor area and presupplementary motor area: targets of basal ganglia and cerebellar output. J. Neurosci. 2007, 27, p. 10659-10673.
2. Banks, P.W., Isham, E.A. We infer rather than perceive the moment we decided to act. Psychol. Sci. 2009, 20, p. 17-21.
3. Brass, M., Haggard, P. To do or not to do: the neural signature of self control. J. Neurosci. 2007, 27, p. 9141-9145.
4. Brass, M., Haggard, P. The hidden side of intentional action: the role of the anterior insular cortex. Brain Struct. Funct. 2010, 214, p. 603-610.
5. Brembs, B. The importance of being active. J. Neurogenet. 2008, 23, p. 120-126.
6. Brembs, B. Toward scientific concept of free will as a biological trait: spontaneous actions and decision-making in invertebrates. Proc. R. Soc. B. 2011, 278, p. 930-939.
7. Custers, R., Aarts, H. The unconscious will: how the pursuit of goals operates outside of conscious awareness. Science 2010, 329, p. 47-50.
8. Dobzhansky, T. Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 1973, 35, p. 125-129. Dostupné též na http://people.delphiforums.com/lordorman/light.htm.
9. Dennett, D. Consciousness explained. London: The Penguin Press, 1991.
10. Dennett, D., Kinsbourne, M. Time and the observer. Behav. Brain Sci. 1992, 15, p. 183-274.
11. Domenici, P., Booth, D., Blagburn, J.M. et al. Cockroaches keep predators guessing by using preferred escapes trajectories. Curr. Biol. 2008, 18, p. 1792-1796.
12. Doyle, R.O. Free will: it´s normal biological trait, not a gift or a mystery. Nature 2009, 459, p. 1052.
13. Doyle, B. Jamesian free will. The two stage-model of William James. William James Studies 2010, 5, p. 1-28.
14. Wikipedia. Free will [on-line]. Dostupné na http://en.wikipedia.org/wiki/Free_will.
15. Friesen, W.O., Kristan, W.B. Leech locomotion: swimming, crawling, and decision. Curr. Opin. Neurobiol. 2007, 17, p. 704 – 711.
16. Haggard, P., Eimer, M. On the relation between brain potentials and the awareness of voluntary movement. Exp. Brain Res. 1999, 126, p. 128-133.
17. Haggard, P. Human volition: towards a neuroscience of will. Nature Reviews Neuroscience 2008, 9, p. 934-946.
18. Hallettt, M. Volitional control of movement: the fysiology of free will. Clin. Neurophysiol. 2007, 118, p. 1179-1192.
19. Heisenberg, M. Is free will an illusion? Nature 2009; 459, p. 164-165.
20. Hume, D. A treatise on human nature (1739) [on-line]. Dostupné na http://oll.libertyfund. org/index.php?option=com_staticxt&staticfile=show.php%3Ftitle=342&Itemid=28.
21. Koukolík, F. Funkční systémy lidského mozku. Galén, Praha 2011(v tisku)
22. Kristan, W.B. Neuronal desicion-making circuits. Curr. Biol. 2008, 18, R929-R932.
23. Kühn, S., Brass, M. Retrospective construction of the judgement of free choice. Conscious. Cogn. 2009, 18, p. 12-21.
24. Libet, B., Gleason, C.A., Wright, W. et al. Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential). The unconscious initiation of freely voluntary act. Brain 1983, 106, p. 623-642.
25. London, M., Roth, A., Beeren, L. et al. Sensitivity to perturbation in vivo implies high noise and suggest rate coding in cortex. Nature 2010, 466, p. 123-127.
26. Nachev, P., Rees, G., Parton, A. et al. Volition and conflict in human medial frontal cortex. Curr. Biol. 2005, 15, p. 122-128.
27. Pessiglione, M., Schmidt, L., Draganski, B. et al. How the brain translates money into force: a neuroimaging study of subliminal motivation. Science 2007, 316, p. 904-906.
28. Searle, J. Minds, brains, and science. Cambridge, MA: Harvard University Press, 1984.
29. Shibasaki, H., Hallett, M. What is the Bereitschaftspotential? Clin. Neurophysiol. 2006, 117, p. 2341-2356.
30. Soon, Ch. S., Brass, M., Heinze, H-J. et al. Unconscious determinants of free decisions in the human brain. Nat. Neurosci. 2008, 11, p. 543-545.
31. Wegner, D.M., Wheatley, T. Apparent mental causation. Sources of the experience of will. Am. Psychol. 1999, 54, p. 480-492.
32.Wegner, D.M. The illusion of conscious will. Cambridge, MA: MIT Press, 2003.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2011 Issue 6
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
Most read in this issue
- Historie a současnost chirurgické léčby TBC plic
- Zánětlivá bolest zad – kdy myslet na spondylartritídu?
- Vazovagální synkopa na kardiologicky profilovaném pracovišti
- Problematika výživy u onkologicky nemocných pacientů