
Základy kognitivní, afektivní a sociální neurovědy
IX. Altruismus
Basics of cognitive, affective and social neuroscience: IX. Altruism
- Hamilton´s kin selection (inclusive fitness),
- Triver’s and Maynard-Smith´s reciprocal altruism
are classical theories explaining biological altruism.
Human altruism is unique in the animal world. It has phylogenetic and ontogenetic roots. Human altruism is based on strong reciprocity – a combination of altruistic punishment and rewarding. Simon’s elegant theory of social selection and succesful altruism explains human altruism in terms of docility and bounded rationality.
- Inequity aversion,
- reciprocal fairness, and
- reputation seeking
as explaining theories are investigated in economic games.
Human altruism has neurobiological correlates evident in experiments making use of functional imaging and molecular genetic methods. Batson’s interesting interpretation explains human altruism as evolution of mammalian and human parental nurturance and empathy.
Key words:
human altruism, evolution, neurobiological correlates, Batson’s interpretation.
Authors:
F. Koukolík
Authors‘ workplace:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
Published in:
Prakt. Lék. 2011; 91(9): 507-511
Category:
Editorial
Overview
Klasické teorie vysvětlující biologický altruismus jsou:
- Hamiltonova teorie příbuzenského výběru (inkluzivní zdatnosti),
- Triversova a Maynardova-Smithova teorie recipročního altruismu.
Lidský altruismus je v živočišné říši unikátní, má fylogenetické i ontogenetické kořeny. Je založen na silné reciprocitě – kombinaci altruistického trestání a odměňování.
Simonova elegantní teorie sociální selekce a úspěšného altruismu vykládá altruismus v pojmech docility a omezené racionality. V ekonomických hrách se coby vysvětlující teorie testují averze vůči:
- nerovnosti,
- reciproční poctivost, a
- vyhledávání reputace.
Lidský altruismus má neurobiologické koreláty patrné v experimentech užívajících funkční zobrazovací i molekulárně genetické metody. Zajímavá Batsonova interpretace vysvětluje lidský altruismus jako vývoj savčího a lidského rodičovského pečování a empatie.
Klíčová slova:
lidský altruismus, evoluce, neurobiologické koreláty, Batsonova interpretace.
Úvod
Problematiku altruismu lze oddělovat od otázek kooperace, obecněji od problematiky sociálního rozhodování (neboli neuroekonomického), jen didakticky (20). S ohledem na rozsah tématu se tato část věnuje altruismu. Kooperaci a sociálnímu rozhodování se budou věnovat následující statě.
Organismus se chová altruisticky, jestliže ho jeho chování něco stojí a přitom prospívá jiným organismům. Náklady a prospěch (cost/benefit) biologicky altruistického chování se měří jako reprodukční zdatnost – počet očekávaných potomků. Chová-li se tedy v biologickém slova smyslu organismus altruisticky, snižuje možný počet vlastních potomků, čímž zvyšuje možný počet potomků druhých organismů. Biologický altruismus není podložen vědomým rozhodnutím o pomoci. Vyskytuje se mezi příslušníky živočišných druhů, u nichž se existence vědomí nepředpokládá. Altruisticky se mohou chovat pravděpodobně všechny podoby života (13).
Darwin (7) si byl vědom, že altruismus je pro evoluci přírodním výběrem problém. Znepokojoval ho hlavně eusociální hmyz, například mravenci nebo termiti. Sterilní dělnice sociálně žijícího hmyzu mají nulovou reprodukční zdatnost, „obětovaly ji“ ve prospěch jiných příslušníků druhu. Přírodní výběr by měl altruismus vyřadit.
Biologický altruismus začaly vysvětlovat dvě klasické teorie
- Hamiltonova teorie příbuzenského výběru (kin selection), respektive teorie inkluzivní zdatnosti (inclusive fitness; (17)
- Triversova (31) a Maynardova-Smithova (24) teorie recipročního altruismu (reciprocal altruism).
Tyto teorie oslabily platnost starší teorie vysvětlující biologický altruismus u lidí skupinovou selekcí (group selection) (7), o jejímž významu pro biologickou evoluci, na rozdíl od Konrada Lorenze, zakladatelé neodarwinismu, R.A. Fisher, J.B.S. Haldane a S. Wright, pochybovali.
Kontroverze v tomto směru trvá, protože někteří autoři považují teorii příbuzenského výběru a evoluční teorii her (evolutionary game theory) za speciální případy skupinové selekce (30). V současnosti teorie skupinové selekce opět ožívá (24, 25).
Základní myšlenka teorie příbuzenského výběru předpokládá existenci genu (nebo genů), jenž svého nositele nutí chovat se k druhým jedincům altruisticky, například sdílet potravu. Organismy, které takový gen nemají, se chovají sobecky, potravu si nechají pro sebe.
Altruisté jsou tedy v nevýhodě.
Jestliže jsou altruisté schopni rozlišovat, s kým budou sdílet potravu, situace se mění. Mohou ji sdílet jen se svými příbuznými. Z toho plyne, že altruista nesoucí kopii altruistického genu bude sdílet potravu s jedincem, jenž pravděpodobně rovněž ponese kopie tohoto genu. Míra pravděpodobnosti záleží na stupni příbuznosti. Altruistický gen snižuje biologickou zdatnost svého nositele, zvyšuje však biologickou zdatnost jeho příbuzných. Počet kopií altruistického genu v další generaci vzroste, výskyt altruistického chování také.
Hamilton (17) dokázal, že přirozený výběr bude „volit“ altruistický gen při naplnění podmínky pojmenované Hamiltonovo pravidlo
b > c/r
kde
b je zisk (benefit),
c jsou náklady (cost), a
r je koeficient vztahu mezi poskytovatelem (dárcem) altruistického chování a jeho příjemcem, vyjadřující míru pravděpodobnosti, s níž dárce a příjemce sdílejí geny v nějakém příbuzensky identickém locusu.
Dva geny jsou příbuzensky identické, jestliže jsou kopiemi jednoho genu společného předka. U druhů, které se reprodukují sexuálně, je r biologických sourozenců 0,5, rodiče a dítěte rovněž 0,5, prarodiče a vnoučata 0,25, bratrance a sestřenice 0,125, atd. Čím je hodnota r vyšší, tím je vyšší pravděpodobnost, že příjemce altruistického chování bude sdílet „gen altruismu“.
Předpověď teorie příbuzenské selekce říká, že se živočiši budou altruisticky chovat ke svým příbuzným s vyšší pravděpodobností, než k jiným příslušníkům vlastního druhu. Nadto bude stupeň altruismu tím vyšší, čím bližší je příbuzenství.
Altruistické chování snižující jedincovu osobní biologickou zdatnost zvyšuje jeho inkluzivní zdatnost definovanou jako osobní zdatnost, k níž se přičítá suma vážených vlivů na zdatnost každého dalšího organismu populace. Váhu vlivu určuje koeficient příbuzenství r. Přírodní výběr tedy bude tímto mechanismem zvyšovat inkluzivní zdatnost příslušníků dané populace. Přitom nejde o přísně deterministický vliv genu coby „příčiny“ nějakého druhu chování, ale o míru pravděpodobnosti, se kterou se bude nositel daného genu chovat: být nositelem „genu pro altruismus“ v tomto modelu znamená pouze vyšší pravděpodobnost altruistického chování.
Teorie příbuzenského výběru nezamítá možnost, že se mnohé druhy chování u živočichů přenášejí negenetickými mechanismy, například imitací a sociálním učením (1).
Hamilton v pozdějších publikacích zdůrazňoval, že jeho původní koeficient příbuzenské blízkosti je nutné nahradit obecnějším korelačním koeficientem určujícím pravděpodobnost, se kterou poskytovatel a příjemce sdílejí geny ať už na příbuzenském, nebo nepříbuzenském základu (18).
Altruismus se v přírodě netýká výlučně příbuzných. Jedno z vysvětlení altruistického chování vůči nepříbuzným členům druhu poskytuje teorie recipročního altruismu (31).
Podle této teorie může být altruistické chování vůči nepříbuzným prospěšné, jestliže lze v budoucnosti očekávat nějakou podobu „splátky dluhu“. „Náklady“ na altruistické chování odpovídají míře pravděpodobnosti, se kterou se „investice“ vrátí.
Reciproční altruismus nevyžaduje, aby daní jedinci byli nutně příslušníci stejného druhu. Nutná je však více než jediná interakce, kromě toho i schopnost v budoucnosti poznat jedince, se kterým byl poskytovatel altruistické péče v interakci a schopnost rozlišit a potrestat parazity.
Lidský altruismus
Vysoká míra altruismu lidský druh od ostatních druhů života odlišuje. Jak v současných společnostech lovců - sběračů, tak ve složitých moderních společnostech existuje síť směnných vztahů, sdílení potravy, dělba práce a kolektivní vedení války. Nejde však o zcela výlučnou vlastnost našeho druhu, byť tomu nasvědčovaly například dosavadní studie pravých šimpanzů (Pan troglodytes verus). Ty však pocházejí z pozorování jejich chování v zajetí. Šimpanzi ve volné přírodě však sdílí potravu, tvoří koalice, kooperativně loví a hlídají hranice svého území, odkud pořádají nájezdy na území sousedících skupin.
Boesch et al. (5) popsal 18 případů adopce osiřelých mláďat šimpanzi žijícími v Národním parku Taï na Pobřeží slonoviny. V polovině případů adoptovali i samci, kteří až na jedinou výjimku nebyli otci mláděte. Neplatí tedy, že „šimpanzi jsou lhostejní k blahu členů skupiny, s nimiž nemají příbuzenský vztah“ (26).
Pro evoluční kořeny svědčí také ontogeneze altruistického chování. Děti ve věku 14–18 měsíců pomáhají druhým jedincům dosáhnout cíl, například získat předmět, který je mimo jejich dosah, nebo otevřít skřínku. Chovají se tak, aniž by za to byly odměňovány dospělým člověkem. Jestliže u dětí starších a dospělých lidí jde při těchto interakcích o reciprocitu a reputaci, v tomto věku se o ně patrně nejedná (32).
Altruistické trestání
Sledujeme-li v běžném životě člověka, který za nějakou cenu pomáhá druhému člověku, nelze obvykle určit příčiny takového chování. Pomáhajícímu může jít o reputaci, může očekávat, že mu člověk, jemuž pomáhá, v budoucnosti pomůže také, může se jednat o pomoc zcela nezištnou. Z tohoto důvodu se při studiu altruismu experimentálně užívají ekonomické hry (11–15).
Příkladem je hra Trestání třetí stranou (third-party punishment game), která je pro tři subjekty A, B, C.
Subjekt A dostane 100 měnových jednotek (MU), B nedostane nic, C dostane 50 MU. A je v diktátorské roli, přidělí „chudému“ B podíl dle vlastní vůle. Jakmile zjistí C výši přesunuté částky, může se, jestliže má pocit, že byl A sobecký, rozhodnout o jeho potrestání, jenže to není zdarma. Za každé 3 MU, které hráč C strhne hráči A, musí hráč C zaplatit 1 MU. Hra je jednokolová, anonymní, takže reputaci nikdo nezískává.
Jestliže by byl hráč C, třetí strana, „ekonomicky racionální“, tedy sobecký, neinvestoval by do trestu nic. V experimentu však 55 % hráčů C (N22) trestalo hráče A, jakmile A přesunul méně než 50 %. Čím byla přesunutá částka nižší, tím byl trest tvrdší. Nadto 70–80 % hráčů B očekávalo, že třetí strana bude trestat, stejně očekávala, že výše trestu bude úměrná klesání přesunuté částky. Trestání za porušenou sociální normu – očekávánou normou je zde přesun poloviční částky – za cenu vlastní oběti je altruistické trestání.
Třetí strany altruisticky trestají podrazáky i ve hře Vězňovo dilema v případech, kdy podrazákův protihráč kooperoval (12).
Altruistické odměňování
Existuje hra pro hráče A, B. Oba dostanou 10 MU. Mohou si ponechat celou částku, mohou však druhé straně poslat libovolný podíl z toho co mají, nebo celou částku. Experimentátor jakoukoli poslanou částku zdvojí. Optimální by tedy bylo, kdyby oba hráči poslali celou částku, v tom případě by oba hráči měli 20 MU. Sobecký hráč si nechává co nejvíc pro sebe, očekává, že mu druhá strana co nejvíc pošle. V případě, že by druhá strana poslala celou svou částku, nezbude jí nic, sobecký, neboli „ekonomicky racionální“ hráč má 30 MU.
Altruistické odměňování se testuje tak, že hráč A posílá podíl nebo celou svou částku první. B je informován kolik A poslal. B se rozhodne, kolik pošle hráči A. Sobecký hráč B by si vše nechal a neposílal by zpátky nic.
Ve skutečnosti však víc než polovina hráčů B peníze zpět pošle, někdy vrácená částka převýší transfer hráče A (4).
Silná reciprocita
Silná reciprocita je kombinace altruistického odměňování, které charakterizuje predisposice odměňovat jiné členy skupiny za kooperaci a za chování dodržujícím normy, spjatá s altruistickým trestáním, což je tendence trestat druhé členy skupiny za porušení norem, a to i za cenu vlastní oběti. Lidé se silnou reciprocitou nesou „náklady“ na altruistické odměňování a trestání, i když z těchto druhů chování nemají žádný ekonomický zisk. Naproti tomu reciproční altruisté biologické literatury odměňují a trestají jen tehdy, jestliže to je v jejich dlouhodobém osobním zájmu (self-interest).
Silná reciprocita je tedy mohutným podnětem pro kooperaci jak v případě interakcí, které se neopakují, tak v případě, že chybí zisky podmíněné reputací, neboť lidé se silnou reciprocitou odměňují lidi, kteří kooperují a trestají lidi, kteří podrážejí.
Klíčem k pochopení lidské kooperace jsou tedy interakce mezi lidmi se silnou reciprocitou a lidmi sobeckými (11).
Chování označované jako nákladná signalizace je podle stejnojmenné teorie (costly signalling theory)
- přínosem pro jiné jedince, kteří takové chování pozorují,
- může být pro signalizujícího jedince nákladné,
- odpovídá biologické zdatnosti signalizujícího jedince.
Může být podnětem recipročně altruistického chování a být výhodné pro volbu sexuálního partnera (16).
Teorie silné reciprocity se neobešla bez vážných námitek, o nichž se zmíním v následující stati o kooperaci.
Simonova teorie sociální selekce a úspěšného altruismu
Zajímavou teorii sociální selekce a úspěšného altruismu formuloval Herbert Simon (27), nositel Nobelovy ceny za ekonomii (1978). Altruismus definuje stejně jako předešlí autoři jako chování zvyšující biologickou zdatnost dalších členů skupiny na vrub zdatnosti vlastní. Zdatností rozumí očekávaný počet potomků.
Pro pochopení altruismu v lidských skupinách je klíčové, zvláště s ohledem na dlouhou dobu nutnou k výchově dětí, sociální učení.
Sociální učení je získávání vědomostí a dovedností od druhých členů skupiny, respektive společnosti. Má dvě stránky:
- jednak zajišťuje znalosti a dovednosti užitečné pro interakce s prostředím,
- jednak zajišťuje učení cílům, hodnotám a postojům vyjadřovaným v chování a řeči dalších členů skupiny.
První stránku nazývá Simon „dovednosti“, druhou stránku „správné chování“.
Učení obou typů přispívá ke zdatnosti jedince. Jedinci se učí úměrně své docilitě, vnímavosti, se kterou přijímají informace, které se učí.
Míra docility je u různých lidí různá, což je částečně podmíněno geneticky. Kromě toho jsou lidé různě inteligentní a jejich motivace je rovněž vzájemně odlišná. Docilní jedinci se učí a věří tomu, čemu jsou učeni, aniž by příliš zkoumali, do jaké míry přispívají naučené informace jejich osobní zdatnosti.
Kromě docility přispívá této skutečnosti omezená racionalita.
Lidská racionalita je nutně omezená díky složitosti světa. Lidé věří ohromnému množství fakt a souvislostí, jimž byli naučeni prostě proto, že nemají možnost si je nezávisle ověřovat.
Přijetí sociálních norem kromě docility napomáhá pocit viny a hanby. Pravděpodobně jsou nezávislé na docilitě. Pocit viny je zvláště významný, protože je efektivní nezávisle na tom, zda byla nonkonformita jedince odhalena, či nikoli. Simonův model dokazuje, že docilní jedinci mají ve společnosti díky své docilitě větší množství potomků než jedinci sobečtí.
Některé druhy chování, byť nákladné pro jedince, prospívají skupině jako celku – například sebeobětování ve válečném konfliktu. Společnost, která těmto druhům chování učí své docilní jedince, poroste rychleji než společnost, která je takovému chování neučí. Docilní jedinec užívající výhody své docility přijme jako součást „správného“ chování altruistické chování ve vztahu ke své skupině. S ohledem na omezenou racionalitu nebude docilní jedinec vždy schopen rozlišit sociálně předepsané chování přispívající zdatnosti od altruistického chování. Docilita bude omezovat schopnost nezávislého vyhodnocení otázky, jak přispívá chování ke zdatnosti.
Pocit viny a hanby budou chování chápané jako altruistické posilovat. Docilní jedinec ponese náklady altruismu. Jestliže nebudou požadavky, které společnost ve věci altruismu vynakládá na docilní jedince vyšší než zisk, jenž jim docilita poskytuje v podobě znalostí a dovedností, bude zdatnost docilních jedinců převyšovat zdatnost jedinců sobeckých.
Je nutné mít na mysli, že jen část altruistova chování snižuje jeho zdatnost. Ne všechno „správné“ chování znamená oběť. Altruismus je v tomto pojetí vedlejším výtvorem docility. Kdyby byla docilita svobodně volenou vlastností, mohl by se altruismus, jenž je jejím výtvorem, jmenovat “osvícené sobectví“.
Docilita, přinejmenším její genetická složka, je však lidem dána. Díky omezené racionalitě docilní jedinec nezvládá učení, které by zvýšilo jeho osobní zdatnost, aniž by se současně nenaučil altruistickému chování, které mu ji snižuje.
Averze vůči nerovnosti
Averze vůči nerovnosti (inequity aversion) (6) je skupina teorií vykládající altruistické chování a kooperaci motivací k ekonomickému zisku, která je současně doprovázena nechutí k nerovnosti. Lidé ji zjišťují srovnáváním s nějakou referenční skupinou, například lékaři se porovnávají s jinými lékaři. Jedinci, kteří jsou na tom lépe, než referenční skupina, mohou (ale nemusí) mít pocit viny, jedinci, kteří na to jsou hůř, mohou (ale nemusí) závidět.
Lidé, kteří mají averzi vůči nerovnosti, kooperují podmíněně tehdy, když kooperuje jejich oponent.
Reciproční poctivost
Teorie reciproční poctivosti (reciprocal fairness), další teoretické vysvětlování altruismu, vychází z předpokladu, že cílem jedince vybaveného silným smyslem pro reciprocitu je odměna poctivého a potrestání nepoctivého „protihráče“ (23). Složitější podoba této teorie má za to, že lidé odhadují motivaci „protihráčů“, odměňují poctivé a trestají nepoctivé záměry, které vyhodnocují na základě možných alternativ.
Záměr totiž může záležet na konkrétní situaci: i poctivý člověk může mít někdy nepoctivý záměr (9).
Evoluční kořeny
Jestliže se averze vůči nerovnosti a reciproční poctivost považují za bezprostřední (proximate) „příčiny“ altruismu, za příčiny základní (ultimate), tedy za evoluční kořeny se považují
- reciproční altruismus, a
- získávání dobrého jména, neboli reputace (reputation seeking).
Reciproční altruismus vystihuje hra Vězňovo dilema (tab. 1) ve vícekolové podobě.

V interakcích dvou osob lidé spolupracují víc, jestliže jsou pravděpodobné další interakce. Model vysvětluje kooperaci i v malých geneticky spřízněných skupinách, nicméně dobře nevysvětluje interakce ve větších skupinách geneticky nespřízněných lidí.
Lidé získávají dobré jméno, neboli reputaci, jestliže pomohou nespřízněnému jedinci a vejde-li to do skupinového povědomí. Tím roste pravděpodobnost, že v budoucnosti někdo pomůže i jim, ale ani tento model není bez problémů (16, 22).
Další možností je koevoluce genů a kultury, jejíž problematika, podobně jako teorie skupinové selekce vyžaduje samostatnou stať.
Neurobiologie
Neuronální podklady altruistického trestání v průběhu ekonomické hry pro dva hráče A, B vyšetřoval de Quervain et al. (10).
Předpokladem experimentu byla kooperace a spravedlivost přikazující hráči B, aby vrátil polovinu získané částky.
Úvaha zněla: jestliže B podrazil, A bude situaci prožívat jako porušení sociální normy, což vyvolá potřebu hráče B potrestat. Hráč A měl možnost udělit hráčovi B až 20 trestných bodů. Rozhodnutí bez udělení trestných bodů se chápalo jako symbolické trestání, rozhodnutí s jejich udělením jako efektivní trestání. Jeden trestný bod znamenal ztrátu 1 MU pro hráče A, 2 MU pro hráče B. Potrestání tedy bylo možné za cenu vlastní ztráty. Na rozhodnutí, zda hráče B potrestá a kolik mu udělí trestných bodů, měl hráč A jednu minutu poté, co se dozvěděl, jak se hráč B rozhodl.
V průběhu této jedné minuty byl mozek hráče A vyšetřován PET.
Efektivní trestání v porovnání se symbolickým trestáním aktivovalo dorzální striatum (obr. 1). Tato oblast se podílí na zpracovávání odměny, která je důsledkem zacílení akce. Jedinci s vyšší mírou aktivace udělovali vyšší tresty. Lidi tedy uspokojuje možnost potrestat narušitele sociální normy. Aktivace dorzálního striata zřejmě odpovídá anticipaci uspokojení z možnosti potrestat podrazáka. Zvýšená aktivace dorzálního striata byla zjištěna i v průběhu anticipace non-sociální odměny, například příjemné chuti, stejně jako pouhého příjmu peněžní částky.
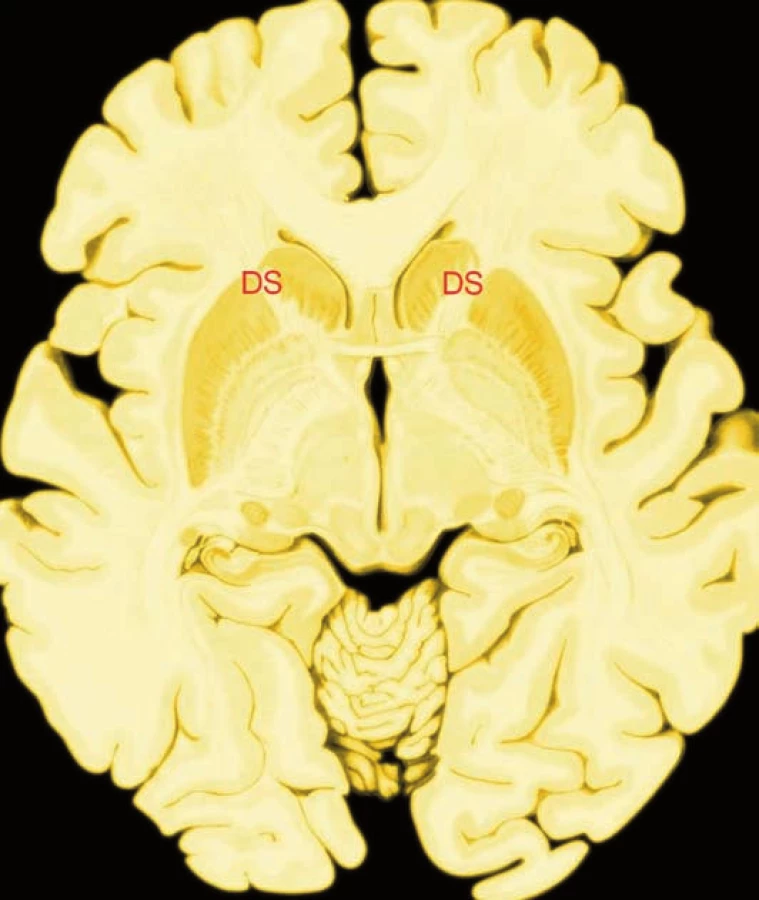
King-Casas et al. (19) na tento experiment navázal testováním neuronální odpovědi dobrovolníků v průběhu vícekolové ekonomické hry pro dva hráče. Zjistil, že reciprocita vyjadřovaná jedním hráčem významně předpovídá budoucí důvěru vyjadřovanou jejich partnerem. I tuto behaviorální odpověď koreluje aktivita dorzálního striata. Míra odpovědi dorzálního striata přitom koreluje se „záměrem důvěřovat“ v dalším kole hry.
Vrchol aktivity striata odpovídající „záměru důvěřovat“ se objevuje 14 sekund poté, co vejde ve známost protihráčova reputace.
Kromě striata se na zpracovávání altruistické odpovědi podílí i aktivita mesencefalických dopaminergních neuronů, které vysílají vlákna do ventrálního striata a ventromediální prefrontální kůry. Rilling et al (29) doložil zvýšenou aktivitu obou oblastí při reciproční altruistické odpovědi, zatímco při opaku aktivita obou oblastí klesala.
Mesolimbický dopaminergní systém tedy „vypočítává“ omyl predikce reciprocity altruistické akce. Jde o druh učení umožňujícího vyhledávat jedince, kteří altruistický čin opětují, a vyhýbat se těm, kteří ho neopětují.
Poměrně častou afektivní a kognitivní reakcí je pocit altruistické viny. Lze se s ním setkat například u lidí, kteří přežijí katastrofickou událost a mají pocit viny vůči těm, kdo ji nepřežili, případně vůči jejich pozůstalým, přestože vědí, že se jednalo o náhodnou událost.
Pocit altruistické viny se odlišuje od pocitu viny deontologické, který se objevuje při narušení niterných hodnot, příkladem je pocit viny katolicky věřící ženy, která absolvovala pohlavní styk před svatbou.
Vyšetření funkční magnetickou rezonancí doložilo odlišnou aktivitu mozku v průběhu pocitu obou typů viny: deontologická vina byla spjata s vyšší aktivací předních úseků gyrus cinguli oboustranně, zatímco pocit altruistické viny doprovázela vyšší aktivita levého mediálního frontálního závitu (2).
Neuropeptidem, který se podílí na regulaci altruistického chování v experimentálním konfliktu mezi lidmi, je oxytocin. Ve třech experimentech užívaly pokusné osoby, muži, oxytocin, nebo placebo (dvojitě slepé uspořádání) v průběhu rozhodování o finančních důsledcích akce
- pro sebe,
- pro členy své skupiny, a
- pro soutěžící cizí skupinu.
Oxytocin zvyšoval vnitroskupinovou důvěru i kooperaci, vůči cizí skupině se chovali muži pod vlivem oxytocinu s agresí označovanou jako defenzivní, nikoli agresí ofenzivní (8).
Prosociální chování jako jsou
- empatie,
- kooperativita, i
- altruismus
mají dědivost (heritability) kolem 0,5.
Podle prvních molekulárně genetických studií korelují míru těchto vlastností polymorfismy genů kódujících receptory neuropeptidů oxytocinu a vazopresinu. Rozptyl vlastností vysvětlených tímto způsobem je však nízký. Z toho plyne, že se podílejí další geneticky podmíněné proměnné.
Reuter et el. (28) proto vyšetřoval polymorfismus COMT Val158Met (gen kódující katechol-o-metyltransferázu, COMT, klíčový enzym metabolismu katecholaminů. COMT se inaktivuje v synaptické štěrbině, hlavně v prefrontální kůře. Jde o polymorfismus jediného nukleotidu, single nucleotide polymorphism, SNP. Záměna kódujícího „písmene“ guaninu za alanin v kodonu 158 genu COMT, který je v proužku q11 chromozomu 22, znamená pro bílkovinný řetěz enzymu vřazení aminokyseliny metioninu na místo aminokyseliny valinu (proto Val1158Met). Výsledkem je pokles aktivity enzymu na třetinu až čtvrtinu.
Lidé s genotypem Val/Val mají aktivitu enzymu nejvyšší, lidé s genotypem Met/Met nejnižší, heterozygoti Val/Met se pohybují uprostřed.
Experimentu se účastnilo 101 mladých zdravých dobrovolníků. Nejprve získávali peníze při dvou namáhavých počítačových pokusech. Pak byli požádáni, aby dle svého uvážení dali nějakou část vydělaných peněz chudému dítěti v rozvojové zemi. Nosiči alespoň jedné alely pro valin dávali přibližně dvojnásobnou částku než lidé, kteří tuto alelu neměli.
Kooperativita a alela pro valin enzymu COMT vysvětlovaly 14,6 % rozptylu altruistického chování měřeného tímto způsobem. Na altruistické chování tedy má vliv i dopaminergní systém.
Odlišný výklad
Odlišný výklad lidského altruismu uveřejnil C. D. Batson (3), sociální psycholog Kansaské univerzity. Klasické biologické teorie považuje pro výklad lidského psychologického altruismu za nedostatečné. Klíčem k altruistickému chování je podle Batsona empatie (přehled 21).
Genetickým podkladem psychologického altruismu je vývoj rodičovské péče. Proběhl ve čtyřech stupních:
- evoluce savců pečujících o mláďata,
- evoluce lidí, (snad i dalších živočišných druhů), schopných chápat jiné lidi jako cítící „agenty“ se záměrným chováním, schopných tedy pochopit i subtilní potřeby druhých lidí,
- vývoj rozvinuté lidské empatie,
- vývoj rozvinutých poznávacích schopností umožňujících rozšířit empatické vciťování, a tím i altruistické chování na lidi, kteří nejsou vlastními potomky.
MUDr. František Koukolík, DrSc.
Oddělení patologie a molekulární medicíny
Národní referenční laboratoř prionových chorob
Fakultní Thomayerova nemocnice
s poliklinikou
Vídeňská 800
140 59 Praha 4 Krč
E-mail: frantisek.koukolik@ftn.cz
Sources
1. Avital, E., Jablonka, E. Animal traditions: behavioural inheritance in evolution. Cambridge: Cambridge University Press 2000.
2. Basile, B., Mancini, F., Macaluso, E. et al. Deontological and altruistic guilt: evidence for distinct neurobiological substrates. Human Brain Mapping 2011, 32, p. 229-239.
3. Batson, C.D. The naked emperor: seeking a more plausible genetic basis for psychological altruism. Economics and Philosophy 2010, 26, p. 149-164.
4. Berg, J., Dickhaut, J., McCabe, K. Trust, reciprocity and social history. Games Econ. Behav. 1995, 10, p. 122-142.
5. Boesch, C., Bolé, C., Eckhardt, N. et al. Altruism in Forest Chimpanzees: The Case of Adoption. PLoS ONE 2010; 5(1): e8901. doi:10.1371/journal.pone.0008901.
6. Bolton, G.E., Ockenfels, E.R.C. A theory of equity, reciprocity, and competition. Am. Econ. Rev. 2000, 90, p. 166-193.
7. Darwin, C. The descent of man and selection in relation to sex. New York: Appleton, 1871.
8. De Dreu, C.K.W., Greer, L.L., Handgraaf, M.J.J. et al. The neuropeptide oxytocin regulates parochial altruism in intergroup conflict among humans. Science 2010, 328, p. 1408-1411.
9. Dufwenberg, M., Kirchsteiger, G. A theory of sequential reciprocity. Games Econ. Behav. 2004, 47, p. 268-298.
10. de Quervain, D.J., Fischbacher, U., Treyer, V. et al. The neural basis of altruistic punishment. Science 2004, 305, p. 1254-1258.
11. Fehr, E., Fischbacher, U. The nature of human altruism. Nature 2003, 425, p. 785-791.
12. Fehr, E., Fischbacher, U. Third-party punishment and social norms. Evol. Hum. Behav. 2004, 25, p. 63-87.
13. Fehr, E., Fischbacher, U. Human altruism – proximate patterns and evolutionary origins. Analyse&Kritik 2005, 27, p. 6-47.
14. Fehr, E., Fischbacher, U. Altruists with green beards. Analyse&Kritik 2005, 27, p. 73-84.
15. Fehr, E. The nature of human altruism. Carl Snyder Memorial Public Lecture. University of California, Santa Barbara 7. 3. 2005.
16. Gintis, H., Smith, E.A., Bowles, S. Costly signalling and cooperation. J. Theor. Biol. 2001, 213, p. 103-119.
17. Hamilton, W.D. The genetical evolution of social behaviour I and II. J. Theor. Biol. 1964, 7, p. 1-32.
18. Hamilton, W.D. Innate social aptitudes in man: an approach from evolutionary genetics. In: R. Fox (Ed.). Biosocial Anthropology. New York: Wiley 1975.
19. King-Casas, B., Tomlin, D., Anen, C. et al. Getting to know you: reputation and trust in a two-person economic exchange. Science 2005, 308, p. 78-83.
20. Koukolík, F. Sociální mozek. Praha: Karolinum, 2006.
21. Koukolík, F. Základy kognitivní, afektivní a sociální neurovědy II. Empatie. Prakt. Lék. 2011, 91, (2) s. 63-67.
22. Leimar, O., Hammerstein, P. Evolution of cooperation through indirect reciprocity. Proc. R. Soc. Lond Series B-Biological Sciences 2001, 268, p. 745-753.
23. Levine, D.K. Modeling altruism and spitefulness in experiments. Rev. Econ. Dyn. 1998, 1, p. 593-622.
24. Maynard-Smith, J. Group selection and kin selection. Nature 1964, 201, p. 1145-1147.
25. Maynard Smith, J. The origin of altruism. Nature 1998, 393, p. 639-640.
26. Silk, J., Brosnan, S., Vonk, J. et al. Chimpanzees are indifferent to the welfare of unrelated group members. Nature 2005, 437, p. 1357-1359.
27. Simon, H.A. A mechanism for social selection and successful altruism. Science 1990, 250, p. 1665-1668.
28. Reuter, M., Frenzel, C., Walter, N.T. et al. Investigating the genetic basis of altruism: the role of the COMT Val158Met polymorphism. Soc. Cogn. Affect. Neurosci. (2010) doi: 10.1093/scan/nsq083.
29. Rilling, J.K., Sanfey, A.G., Aronson, J.A. et al. Opposing BOLD responses to reciprocated and unreciprocated altruism in putative reward pathways. Neuroreport 2004, 15, p. 2539-2543.
30. Sober, E., Wilson, D.S. Unto others: the evolution and psychology of unselfish behavior. Cambridge MA: Harvard University Press, 1998.
31. Trivers, R.L. The evolution of reciprocal altruism. Q. Rev. Biol. 1971, 64, p. 35-57.
32. Warneken, F., Tomasello, M. The roots of human altruism. Br. J. Psychol. 2009, 100, p. 455-471.
Labels
General practitioner for children and adolescents General practitioner for adultsArticle was published in
General Practitioner

2011 Issue 9
- Advances in the Treatment of Myasthenia Gravis on the Horizon
- Hope Awakens with Early Diagnosis of Parkinson's Disease Based on Skin Odor
- Memantine in Dementia Therapy – Current Findings and Possible Future Applications
- Memantine Eases Daily Life for Patients and Caregivers
- Possibilities of Using Metamizole in the Treatment of Acute Primary Headaches
Most read in this issue
- Ortostatická hypotenzia
- Glomus tumor prstu
- Demodikóza
- Interakce alkoholu a jiných látek: komplikovaný problém