
Developmental Link between Sex and Nutrition; Regulates Sex-Specific Mandible Growth via Juvenile Hormone Signaling in Stag Beetles
Sexual dimorphisms in trait expression are widespread among animals and are especially pronounced in ornaments and weapons of sexual selection, which can attain exaggerated sizes. Expression of exaggerated traits is usually male-specific and nutrition sensitive. Consequently, the developmental mechanisms generating sexually dimorphic growth and nutrition-dependent phenotypic plasticity are each likely to regulate the expression of extreme structures. Yet we know little about how either of these mechanisms work, much less how they might interact with each other. We investigated the developmental mechanisms of sex-specific mandible growth in the stag beetle Cyclommatus metallifer, focusing on doublesex gene function and its interaction with juvenile hormone (JH) signaling. doublesex genes encode transcription factors that orchestrate male and female specific trait development, and JH acts as a mediator between nutrition and mandible growth. We found that the Cmdsx gene regulates sex differentiation in the stag beetle. Knockdown of Cmdsx by RNA-interference in both males and females produced intersex phenotypes, indicating a role for Cmdsx in sex-specific trait growth. By combining knockdown of Cmdsx with JH treatment, we showed that female-specific splice variants of Cmdsx contribute to the insensitivity of female mandibles to JH: knockdown of Cmdsx reversed this pattern, so that mandibles in knockdown females were stimulated to grow by JH treatment. In contrast, mandibles in knockdown males retained some sensitivity to JH, though mandibles in these individuals did not attain the full sizes of wild type males. We suggest that moderate JH sensitivity of mandibular cells may be the default developmental state for both sexes, with sex-specific Dsx protein decreasing sensitivity in females, and increasing it in males. This study is the first to demonstrate a causal link between the sex determination and JH signaling pathways, which clearly interact to determine the developmental fates and final sizes of nutrition-dependent secondary-sexual characters.
Published in the journal:
Developmental Link between Sex and Nutrition; Regulates Sex-Specific Mandible Growth via Juvenile Hormone Signaling in Stag Beetles. PLoS Genet 10(1): e32767. doi:10.1371/journal.pgen.1004098
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1004098
Summary
Sexual dimorphisms in trait expression are widespread among animals and are especially pronounced in ornaments and weapons of sexual selection, which can attain exaggerated sizes. Expression of exaggerated traits is usually male-specific and nutrition sensitive. Consequently, the developmental mechanisms generating sexually dimorphic growth and nutrition-dependent phenotypic plasticity are each likely to regulate the expression of extreme structures. Yet we know little about how either of these mechanisms work, much less how they might interact with each other. We investigated the developmental mechanisms of sex-specific mandible growth in the stag beetle Cyclommatus metallifer, focusing on doublesex gene function and its interaction with juvenile hormone (JH) signaling. doublesex genes encode transcription factors that orchestrate male and female specific trait development, and JH acts as a mediator between nutrition and mandible growth. We found that the Cmdsx gene regulates sex differentiation in the stag beetle. Knockdown of Cmdsx by RNA-interference in both males and females produced intersex phenotypes, indicating a role for Cmdsx in sex-specific trait growth. By combining knockdown of Cmdsx with JH treatment, we showed that female-specific splice variants of Cmdsx contribute to the insensitivity of female mandibles to JH: knockdown of Cmdsx reversed this pattern, so that mandibles in knockdown females were stimulated to grow by JH treatment. In contrast, mandibles in knockdown males retained some sensitivity to JH, though mandibles in these individuals did not attain the full sizes of wild type males. We suggest that moderate JH sensitivity of mandibular cells may be the default developmental state for both sexes, with sex-specific Dsx protein decreasing sensitivity in females, and increasing it in males. This study is the first to demonstrate a causal link between the sex determination and JH signaling pathways, which clearly interact to determine the developmental fates and final sizes of nutrition-dependent secondary-sexual characters.
Introduction
The evolution of sex-specific traits in animals has long fascinated biologists. How is growth regulated so that it differs dramatically between males and females? Sexual dimorphisms are widespread across diverse animal taxa and include exaggerated sexually selected traits like the antlers of deer, the enormous clawed chelae of crabs, and the elaborate trains of peacocks [1], [2], [3]. Some of the most striking sexually dimorphic traits are found within insects, such as the horns of rhinoceros beetles and the large mandibles of male stag beetles [3], [4], [5].
Sex-specific exaggerated traits often develop in a condition-dependent manner, so that not all individuals produce the trait even in the same sex [6], [7], [8], [9], [10]. Virtually all of the most extreme ornaments and weapons are also conditionally-expressed; they are exquisitely phenotypically plastic structures, whose growth depends on larval/juvenile access to nutrition [3], [5], [9], [11], [12]. Consequently, developmental mechanisms generating sex-specific trait growth and nutrition-dependent phenotypic plasticity are each likely to regulate the expression of extreme structures of sexual selection. The near universality of sex differences in the nutrition sensitivity of these traits suggests that common developmental mechanisms may be involved. Yet we still know almost nothing about how the processes of sex-specific growth and nutrition-sensitivity interact with each other to generate sexual dimorphism.
Recent studies in model organisms such as the fruit fly, nematode, medaka fish, and mouse, implicate a group of highly conserved proteins known as DM, or DNA binding motif proteins, as major effectors of sexual differentiation (recently reviewed in [13] and [14]. The fruit fly DM domain gene Doublesex (dsx) is conserved in structure and function in all insect species where it has been examined [2], [15], [16], [17], [18], [19], [20], [21], [22]. The dsx gene is transcribed in both sexes, but then differentially spliced to produce a male-specific or a female-specific mRNA (for review see [13]). These alternatively-spliced sex-specific transcripts code for a male (DsxM) or a female-specific (DsxF) protein [13]. Both types of Dsx proteins contain a zinc finger-like DNA binding domain called the DM domain [23], and act as transcriptional regulators responsible for sexual differentiation of tissues during development [1], [2], [24], [25]. For these reasons, dsx is a promising candidate for the regulation of sexual dimorphisms in the weapons of beetles. Indeed, recent published papers on dsx function in dung beetles (Onthophagus taurus and O. sagittarius) and rhinoceros beetles demonstrate a functional role for dsx in sex-specific growth of horns [20], [22], [26].
In stag beetles, many species show strong sexual dimorphism in the size of their mandibles [5], [27], [28], [29], [30], [31], [32]. Males that have access to unlimited amounts of food as larvae develop disproportionately larger mandibles than males with restricted access to food, but more importantly, female mandibles never proliferate to the extent of even poorly-fed, small males (Fig. 1) [5]. Gotoh et al. recently found that nutrition-dependent mandibular growth in stag beetles is mediated by juvenile hormone (JH) in a sex-specific fashion [5]. JH titers were positively correlated with individual nutritional condition, and, in males, high JH titers promoted the growth of mandibles. In contrast, although females had similar levels of JH to males, female mandibles did not respond to high JH. Also, JH treatment did not affect to the growth of mandibles in females. These results indicate a sex-specific response of these traits to nutritional condition via JH [5]. However, other than this intriguing result, the mechanisms underlying developmental links between sex-determination, endocrine signaling, and sex-specific trait growth have yet to be characterized for any insects with exaggerated sexual dimorphism in insects.

Here, we investigated the developmental mechanisms of sex-specific mandible growth in the stag beetle Cyclommatus metallifer, focusing on dsx gene function and its interaction with JH signaling. This species was used in previous studies on mandible development [5], [33] and we have recently constructed a transcriptome database for this species (Gotoh et al. in prep). To characterize dsx in Cyclommatus metallifer, the full length C. metallifer dsx (hereafter Cmdsx) transcript was obtained by degenerate PCR and subsequent Rapid Amplification of cDNA Ends PCR (RACE-PCR). Expression analyses of Cmdsx were carried out by Reverse Transcription PCR (RT-PCR) and real-time quantitative PCR (qPCR) to reveal the spatio-temporal expression pattern and sex-specificity of the Cmdsx transcripts during the prepupal period, which is known to be the critical period when mandibular tissues proliferate to their final adult size (Fig. 2) [5]. The function of dsx during sex-specific morphogenesis was investigated by gene knockdown using RNA interference (RNAi) against the Cmdsx transcripts. In addition to this, to investigate the putative interaction between dsx and endocrine (JH) signaling during mandibular growth, we ectopically applied JH analog to dsxRNAi individuals.

Results
Identification of sex-specific alternative splice variants of Cyclommatus metallifer dsx
The full-length Cmdsx transcript was obtained by degenerate PCR and subsequent RACE-PCR (Fig. 3A). Four distinct splice variants (A, B, C and D isoforms) were identified, which contain the highly conserved DM domain and encode protein sequences with high sequence similarity to known insect Dsx proteins (Fig. S1, [34]). Protein sequence similarity of the stag beetle isoforms with that of the recently reported doublesex gene of Onthophagus taurus (Scarabaeidae, Coleoptera; [20]), and RT-PCR expression analyses show that CmdsxA and CmdsxB are male-specific, while isoforms CmdsxC and CmdsxD are female-specific (Fig. 3B).

There are several differences in structure among the four splice variants of the Cmdsx transcript. First, a large exon (exon 4) containing primarily noncoding sequence and showing no similarity to dsx exons in any other insects, only occurs in splice variant A (Fig. 3A). In addition, CmdsxA contains neither exon 8 nor exon 9 (found in variants B, C, and D). Cmdsx variants C and D are similar to each other overall, with the exception of exon 6 which is only found in CmdsxC (Fig. 3A). An important difference between the predicted isoforms is the absence of 14 amino acid residues at the 3′-end of the conserved dsx dimer domain in the A and B isoforms (Fig. S1); this difference in the dsx dimer domain was also reported in the doublesex gene of Onthophagus taurus [20]. CmDsxB and the male-specific Dsx isoform of O. taurus (OtDsxM) also share a 25 amino acid sequence at their 3′ end (Fig. S1). Also CmDsxC and CmDsxD had similar 3′ end sequences to the female-specific Dsx isoforms of O. taurus (OtDsxF1 and OtDsxF2), respectively (Fig. S1).
Sex- and tissue-specific expression of dsx during mandible growth
Expression patterns of exons of the Cmdsx transcript were examined in developing mandibles of both sexes in detail by real-time qRT-PCR during different stages of prepupal development (Fig. 3C) Tissue-specific expression and nutrition-dependent expression were examined by measuring the expression level of exon 1, which represents the total Cmdsx isoform expression since it is shared by all splice variants. Also, expression patterns of exons 4, 6, 8 and 9 were examined in order to characterize the sex-specific usage of these exons.
Mandibles are sexually dimorphic (e.g. males have disproportionately large mandibles, Fig. 1B) and were expected to show high Cmdsx expression. Maxillae, on the other hand, are not dramatically different in the two sexes (e.g. maxillae show isometric allometry in both sexes, Fig. 1C) and we expected lower levels of expression of Cmdsx. As predicted, expression of exon 1 was higher in mandibles than in maxillae in both males and females, especially during prepupal Stages 1 and 2 (Fig. 3C). Exon 1 was expressed at its highest level in male mandibles during prepupal Stage 2, but peaked later during (Stage 3) in female mandibles (Fig. 3C). No differences in expression of exon 1 were detected in Stage 4 of prepupal development in either trait (Fig. 3C).
Large and small males have different nutritional histories and undergo different amounts of mandible growth. However, expression levels of Exon 1 were similar for large and small males, during Stages 1, 3 and 4 in both mandibles and maxillae (Fig. 3C). Only during Stage 2 were there significant differences in expression of Exon 1 (Fig. 3C).
Sex-specificity of each exon was examined during Stages 2 and 3, when total Cmdsx expression reached its peak in males and females (Fig. 3C). Expression levels of Exon 4 were five times higher in males than females during Stage 2 (Fig. 3C) and only low levels of expression of this exon were found in both males and females during Stage 3 (Fig. 3C). The female-specific exon 8 was more highly expressed in females during both stages 2 and 3 but especially during Stage 3 (Fig. 3C). Exon 6 and exon 9 were expressed similarly in both males and females during Stage 2, but showed increased expression in females during Stage 3 (Fig. 3C). The increase of expression of exons 6 and 9 in Stage 3 females is expected to correspond to an overall expression peak of all dsx isoforms in females during Stage 3 (Fig. 3A)
Functional analysis of dsx reveals a role in sex-specific mandible growth
Injections of Cmdsx dsRNA reduced Cmdsx transcript abundances by 13–84% in prepupal mandibles, compared with control injections of GFP dsRNA (Fig. S2). The region of the Cmdsx transcript that was targeted extended from exon 1 to exon 3, and was therefore predicted to knockdown all four of the expressed transcripts of Cmdsx in both sexes (Fig. 3A). Knockdown of the Cmdsx gene by RNAi during prepupal development confirmed a significant functional role in the regulation of sex-specific mandible growth in stag beetles. The phenotype of dsxRNAi females was changed to be more male-like in body color, mandible size, foreleg tibial spine number, and genital shape and genital size (Fig. 4A, B, C, D). Significant mandible growth was induced in dsxRNAi females compared with GFPRNAi females (t = 4.509, P = 0.000357, Fig. 4B). In contrast, in dsxRNAi males, mandible growth was dramatically and significantly suppressed (Fig. 4E, F), resulting in more female-like forms. The relationships of body size and mandible size are significantly different between GFPRNAi and dsxRNAi males (F = 19.072, P = 0.0002982). In the range of the observed body size, dsxRNAi males possessed smaller mandibles, and the mandible-size difference became larger as body size increases (Fig. 4F). The intersex phenotypes resulting from the dsxRNAi knockdown extended to the body color of females, transforming them from the black color typical of females to a metallic copper typical of males (Fig. 4A). The number of spines on the tibia also changed. Females typically develop with four or five tibial spines (and males with zero). However, in dsxRNAi females this number decreased from four to fewer (becoming more male-like), and in dsxRNAi males the number of spines increased from zero to four (more female-like) (Fig. 4C, G). Finally, the size and length of the genitalia changed in sex-inappropriate directions in both dsxRNAi females and dsxRNAi males (Fig. 4D, H).

Dsx modulates the response to JH in a sex-specific manner
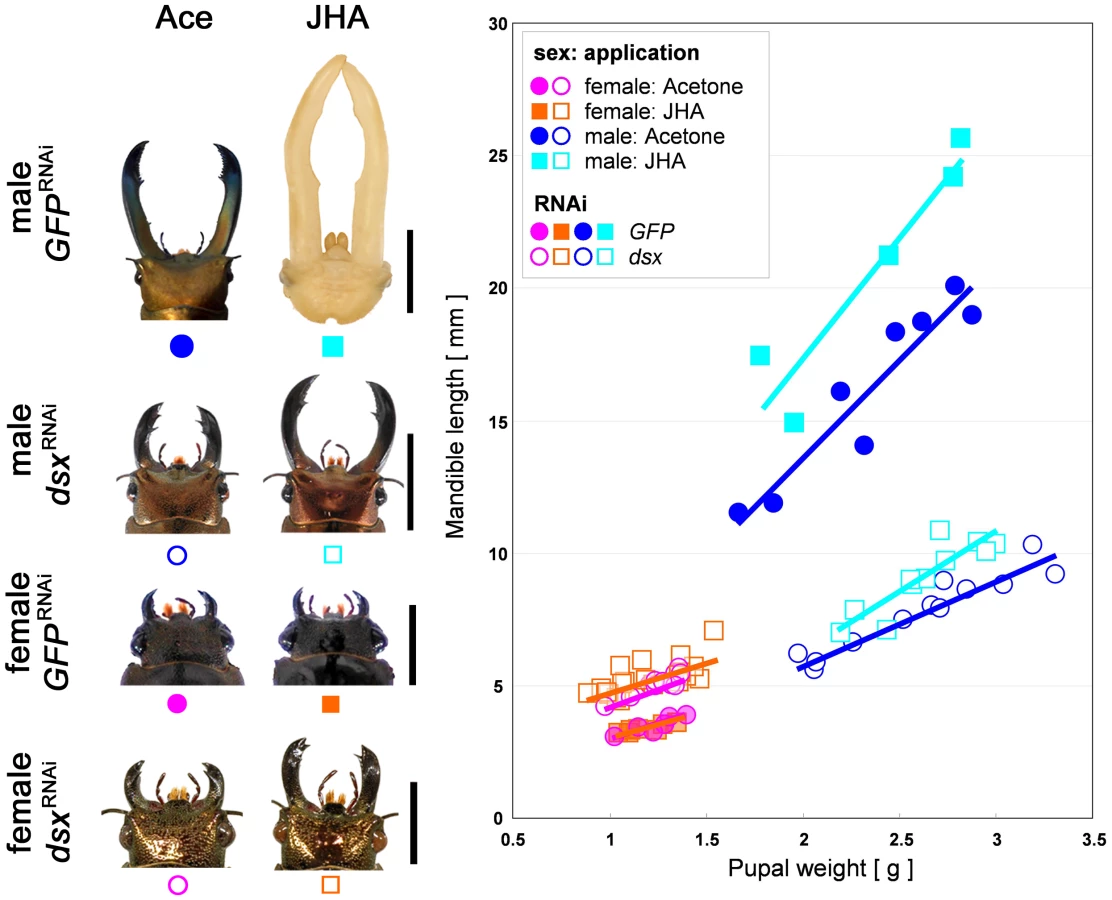
We have previously shown that JH titer during the prepupal period is correlated with adult male body size and mandible size in the stag beetle, and that ectopic application of JH to the prepupal male induces male mandible proliferation [5]. At least part of the exquisite condition-sensitivity of extreme mandible growth appears to involve sensitivity of mandibular tissues to circulating JH. However, we have also shown that mandibular tissues of females do not respond to JH in the same way as males. Mandibles in females did not respond to ectopic JH, despite the fact that females had similar levels of circulating JH to males during this developmental period [5]. This suggests that female mandibular tissues may be insensitive to JH signaling.
We predicted that sex-differences in tissue sensitivity to JH could be caused by action of the sex-determination cascade, specifically by expression of alternative splice variants of Cmdsx. To test for a functional role of Cmdsx in causing sex differences in the sensitivity of mandibular cells to JH, we applied a JH analog (JHA) to RNAi (GFP or dsx) treated males and females (Fig. 5). In control (GFPRNAi) females, JHA application did not induce mandible growth (t = −0.611, P = 0.5549390199, Fig. 5), which corroborates our previously reported result that JHA application does not affect mandibular growth in wild-type females [5]. In contrast, JHA application to dsxRNAi females induced significant growth of mandibles compared with acetone application to dsxRNAi females (t = 2.254, P = 0.0429177662, Fig. 5). Thus, knockdown of Cmdsx caused mandibles of females to behave like those of males. Their growth became sensitive to JH, and therefore should also have become condition-dependent. In control (GFPRNAi) males, JHA application induced significant mandible growth (t = 5.5500, P = 0.0004876470, Fig. 5), corroborating our earlier report that JHA application promotes mandibular growth in wild-type males [5]. For dsxRNAi males, JHA application rescued the defective mandibular phenotype by promoting mandibular growth (ANCOVA, t = 4.918, P = 0.0003321826, Fig. 5), however, the effect of JHA application tended to be decreased in dsxRNAi individuals. We suspect that here, too, the result was to make mandibles in males behave more like those of females. That is, growth of mandibles in knockdown males might be less sensitive to JH than it otherwise would have been.
Discussion
Doublesex and sex-specific trait expression
Spatiotemporal patterns of expression and functional analyses of Cmdsx support the hypothesis that sex-specific growth of exaggerated mandibles in stag beetles is controlled by doublesex. The differences in the 3′ end between male- and female-specific CmDsx are predicted to have important consequences for DNA binding. In Drosophila, it is known that this domain enhances DNA recognition by promoting dimerization of Dsx [35]. Thus, this difference of domain structure in CmDsx suggests differential DNA-binding ability of the predicted male and female proteins. In addition to the differences between sex-specific isoforms, all four isoforms differ from each other in their amino-acid sequence at the 3′ end, raising the possibility that each isoform is deployed differentially in space and time in a sex- and tissue-specific manner. Expression analyses in other body parts and isoform-specific knockdown experiments will be required to confirm this possibility.
Examination of Cmdsx expression in the sexually dimorphic mandibles compared to the sexually monomorphic maxillae revealed that there are differences in expression in a developmental and tissue specific pattern (Fig. 3C). Recent work in Drosophila showed that dsx expression was temporally and spatially restricted to body parts showing sexual dimorphism [36], [37]. In stag beetle mandibles, female-specific Cmdsx transcripts showed their highest levels of expression at the exact stage (Stage 3 of prepupal development) when mandibular cells proliferate maximally in males [5]. Thus, female specific Cmdsx expression coincides precisely with inhibition of mandibular cellular proliferation (Fig. 3C). Expression of male-specific Cmdsx transcripts in mandibles peaked just before this stage, during a period (prepupal Stage 2) when cells in male mandibles are especially sensitive to the growth-promoting effects of JH [5]. Both of these sex- and trait-specific patterns of expression are consistent with isoform-specific regulatory roles for Cmdsx during mandible growth. Considering the results of Cmdsx knockdown, we suggest that male-specific Cmdsx transcripts may promote mandible growth, and female-specific transcripts inhibit mandible growth, in part by enhancing or repressing the sensitivity of mandibular cells to JH.
Doublesex and juvenile hormone signaling
Our results demonstrate, for the first time in any insect, a functional link between Dsx expression and JH signaling. Knockdown animals had significantly altered responses to topical JHA application, compared with control animals (Fig. 5). In females, knockdown of Cmdsx caused mandibles to be sensitive to JHA, where they otherwise would not have been, suggesting that normal expression of female-specific isoforms of CmDsx contributes to insensitivity of female mandibles to JH. Because JH acts to stimulate cell proliferation during this developmental stage, such a mechanism would repress excessive growth of this structure in females. In males, knockdown of Cmdsx combined with topical application of JHA stimulated some mandible growth, but not as much as in control animals with application of JHA. This indicates that male mandibles retained some sensitivity to JH even in their lowered expression of Cmdsx. We suggest that some sensitivity of mandibular cells to JH is the default developmental state for these animals. In normal males, male-specific CmDsx isoforms may increase the sensitivity of mandibular cells to JH, contributing to rapid and disproportionate growth of these exaggerated structures. In this case, knockdown of Cmdsx would remove this extra-sensitivity, restoring mandibular cells to their default state and producing males with large, but not extreme, mandible sizes. Another possibility is that dsx and JH act in parallel to regulate mandible growth in males. If this were the case, then Cmdsx and JH would act independently to stimulate exaggerated growth of male mandibles, and their effects would simply be additive. Thus, crosstalk between dsx and JH in males will need to be investigated in future studies.
Although there have been many previous reports of sex-specific JH actions on secondary-sexual characters in various insect lineages [38], [39], [40], [41], this study is the first to demonstrate a causal link between the JH signaling pathway and the sex determination pathway, which clearly interact to determine the developmental fates of secondary-sexual characters.
The genetic and physiological mechanisms underlying sexually dimorphic traits
A recent study on dsx regulation of sexual dimorphism has also been reported for horned beetles (Onthophagus taurus) [20]. In this study, critical roles of dsx in sex differentiation, including development of sex-specific exaggerated traits, were shown. Horns in O. taurus are dimorphic in two ways: females do not produce horns (sexual dimorphism), and males smaller than a threshold body size produce only rudimentary horns (male dimorphism). Kijimoto et al showed that only large males expressed the male specific isoform of Otdsx (OtdsxM); small males did not [20]. Because body size and horn morphology depend critically on nutrition in this species, the findings of Kijimoto et al raise the possibility that levels of expression of OtdsxM may be sensitive to nutrition, as well as sex [20]. This contrasts with C. metallifer, where horns do not exhibit male dimorphism (all males produce enlarged mandibles) and where we find at best minimal evidence of nutrition-dependent expression of dsx (based on comparisons between large and small males). Expression of Cmdsx in mandibles of large males was at most 1.3 times that of mandibles in small males (during Stage 2; Fig. 3C), which is much smaller than the differences observed for Onthophagus (large males showed approximately 3 times higher expression of OtdsxM than small males). Based on these results we suggest that endocrine pathways sensitive to nutrition may interact with the sex determination pathway both upstream [20] and downstream (our study) of dsx.
Additional developmental factors are likely involved in development of sexually dimorphic traits
The inability of JHA treatment of dsxRNAi males to induce full growth of mandibles may indicate the action of other regulatory pathways for mandible growth. One likely candidate is the insulin-signaling pathway, because this pathway is known to regulate body and organ size in insects in accordance with nutritional conditions [42], [43], [44]. Growing horns in male rhinoceros beetles (Trypoxylus dichotomus) are known to be more sensitive to insulin signals than other metric traits (e.g., wings, genitalia) [3], and Emlen et al. reported sex- and morph- (major vs minor male) specific expression of the insulin receptor (InR) in growing horns of the dung beetle (Onthophagus nigriventris) [10]. It is likely that the enlarged mandibles of male stag beetles will also be sensitive to insulin signaling during their period of growth, and we suspect that Cmdsx may contribute to sex differences in sensitivity to these signals as well (Fig. 6). Future studies such as expression analyses of InR and insulin-like peptides will be needed to examine these additional mechanisms, but already it is clear that a rich interplay between endocrine and sex-determination pathways coordinates the growth of exaggerated sexually-selected and sexually-dimorphic characters.

Materials and Methods
Insect husbandry
Stag beetle adults (Cyclommatus metallifer) were purchased from Hercules-Hercules, Sapporo, Japan. Detail rearing and breeding methods are described in the Supporting Information (Text S1).
Developmental staging of the prepupal period
We defined four developmental stages during the prepupal period based on specific developmental landmarks (Fig. 2). First, the stag beetle final instar larva constructs a pupal cell prior to pupation which marks the border between the end of the larval stage and the onset of the prepupal stage. After pupal cell construction (PCC), the larva undergoes a two-stage gut purge (GP) in which all gut contents are egested from the body. It takes two days from the onset of PCC to the start of the first GP, which is termed ‘Stage 1’ (Fig. 2). The period of time that the first GP continues lasts about 3–4 days and is ‘Stage 2’ (Fig. 2). Overall body weight gradually decreases during stage 2 (Fig. 2). After the first GP is over, the individual remains in a suspended state for 3–5 days during which time the adult structures are proliferating and growing (Stage 3). Stage 4 is a very brief period which corresponds just a few hours prior to pupation when the individual purges all of its remaining gut contents for the second GP and completes metamorphosis into the pupal stage.
Cloning of dsx and reference genes from C. metallifer
Partial transcript sequences of the C. metallifer orthologs for dsx were cloned by degenerate PCR. Three additional transcripts for C. metallifer reference genes for real time qPCR were also cloned by degenerate PCR – glyceraldehyde-3-phosphate dehydrogenase (GAPDH), elongation factor 1 alpha (EF-1a) and ribosomal protein L32 (rpl32). Primer sequences for degenerate PCR are listed in Table S1. Data base searches for homologies were performed using BlastX at the NCBI server (http://blast.ncbi.nlm.nih.gov/Blast.cgi). To further confirm the orthologues, we made multiple alignments of dsx genes including the orthologues from the other insect species, and constructed neighbor-joining trees of protein sequences (for Cmdsx) or mRNA sequences (for GAPDH, EF-1a and rpl32) using ClustalX program [45] (http://www.clustal.org/) (Fig. S3, S4, S5). Confidence was estimated with 1000bootstraps. Detailed cloning procedures are described in the Supporting Information (Text S1).
Identify full-length of dsx gene by using RACE
Rapid amplification of cDNA ends (RACE) -PCR was performed to obtain the full length C. metallifer dsx transcript sequence using the following RACE primers (for 5′-RACE: 5′- CCT GAA CAC GTC GGG AAA AGA CGG CG-3′, for 3′-RACE: CTC GAA GAT TGC CAT AAG CTC CTG GAA AGG-3′) and the SMART RACE cDNA Amplification Kit (Clontech, Palp Alto, CA). The amplified cDNA fragments were subcloned and sequenced as described before. The protein domains were predicted by using CDD (conserved domain database) on NCBI (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi).
RT-PCR and real-time quantitative PCR for Cmdsx expression analysis
The expression patterns of Cmdsx transcripts in small and large male and female prepupae were examined during the period of maximal mouthpart growth with RT-PCR and real-time qPCR. Briefly, individuals were reared under high versus low nutrition conditions that result in small versus large prepupae; details for this can be found in our previous study [5]. See Supporting Information (Text S1 and Table S2) for a detailed description of our methods for RT-PCR and real-time qPCR and primer sequences for real-time PCR.
Knock-down of dsx by RNA interference
Functional analysis of the stag beetle dsx was accomplished by knockdown of the dsx transcript by RNA interference (RNAi) during prepupal development. To silence all Cmdsx transcripts, double-stranded RNA (dsRNA) against a 352 bp region of Cmdsx common to all four splice variants (Fig. 3A) was synthesized. Detailed procedures for dsRNA synthesis are described in the Supporting Information (Text S1). All dsRNA was diluted with 1×PBS. One µg of dsRNA in 5 µl of PBS was injected into the dorsal prothorax of late 3rd instar larvae using a microliter syringe (Hamilton, Reno, NV) under a stereomicroscope. This stage is just prior to the prepupal period and prior to adult mandibular cellular proliferation, so the effect of RNAi was targeted to pupal development (Fig. 2). Individuals that successfully eclosed into adults were used for analyses; these included 7 GFPRNAi females, 12 dsxRNAi females, 8 GFPRNAi males, and 16 dsxRNAi males.
For statistical test of Cmdsx RNAi effect, analysis of covariance (ANCOVA) was performed with body size as a covariate using R 3.0.1 software [46]. The equality of the slopes of regression lines was tested and no significant interaction was detected in female samples (F = 0.2779, P = 0.6058). In male samples, the slopes were significantly different between GFPRNAi and dsxRNAi samples (F = 19.072, P = 0.0002982).
RNAi efficiency was also examined by measurement of Cmdsx expression levels using real-time qPCR in the prepupal mandibles of males and females injected with dsRNA against GFP (control) or Cmdsx. Primers for real-time qPCR were designed to the common region shared by all isoforms (forward primer: 5′-TTC CGC TCT CAT TCA TAA ACGA-3′, reverse primer: 5′-TGC GGA AAA CGG CAA AGT-3′). To prevent overestimation of transcripts by detecting injected dsRNA, we designed the primers to amplify a region that had no overlap with the region used in dsRNA synthesis.
JHA treatment on GFP and dsx RNAi individuals
To investigate the effects of Cmdsx on JH action, we combined a Cmdsx knockdown experiment with ectopic application of the JH analog (JHA). According to previous study [5], we used fenoxycarb for JHA application. First, we injected dsRNA against GFP or Cmdsx into the dorsal thorax of late 3rd instar C. metallifer larvae as described above. Then, when the knockdown experimental prepupae reached stage 2, five µg of the JH analog fenoxycarb diluted in 10 µl of acetone (Wako) was applied to the dorsal thorax according to previous study [5]. The control groups were dsRNA-injected pupae treated with acetone only. Pupal weight and pupal (in males) or adult (in females) mandible length were recorded. Sample sizes of surviving animals with normal, measurable traits are described in Table S3.
To estimate the effect of JHA application on relative mandible size for each of the four RNAi categories (GFPRNAi males, dsxRNAi males, GFPRNAi females, dsxRNAi females), analysis of covariance (ANCOVA) was performed with body size as a covariate using R 3.0.1 software [46]. The equality of the slopes of the regression lines was tested and no significant interaction was detected in all the four RNAi categories (F = 0.6789, P = 0.4313 in GFPRNAi males; F = 2.9354, P = 0.1029 in dsxRNAi males; F = 2.4669, P = 0.1507 in GFPRNAi females, F = 0.6751, P = 0.4185 in dsxRNAi females). Statistical significance of JHA application effects was adjusted for multiple comparisons by using Benjamini & Hochberg method [47].
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. WilliamsTM, CarrollSB (2009) Genetic and molecular insights into the development and evolution of sexual dimorphism. Nat Rev Genet 10: 797–804.
2. GempeT, BeyeM (2011) Function and evolution of sex determination mechanisms, genes and pathways in insects. Bioessays 33: 52–60.
3. EmlenDJ, WarrenIA, JohnsA, DworkinI, LavineLC (2012) A mechanism of extreme growth and reliable signaling in sexually selected ornaments and weapons. Science 337: 860–864.
4. EmlenDJ (2008) The evolution of animal weapons. Annu Rev Ecol Evol Syst 39: 387–413.
5. GotohH, CornetteR, KoshikawaS, OkadaY, LavineLC, et al. (2011) Juvenile hormone regulates extreme mandible growth in male stag beetles. PLoS ONE 6(6): e21139 Available: http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0021139.
6. Andersson M (1994) Sexual Selection. Princeton: Princeton Univ. Press.
7. KnellRJ, FruhaufN, NorrisKA (1999) Conditional expression of a sexually selected trait in the stalk-eyed fly Diasemopsis aethiopica. Ecol Entomol 24: 323–328.
8. EmlenDJ, NijhoutHF (2000) The development and evolution of exaggerated morphologies in insects. Annu Rev Entomol 45: 661–708.
9. CottonS, FowlerK, PomiankowskiA (2004) Do sexual ornaments demonstrate heightened condition-dependent expression as predicted by the handicap hypothesis? Proc R Soc Lond B Biol Sci 271: 771–783.
10. EmlenDJ, SzafranQ, CorleyLS, DworkinI (2006) Insulin signaling and limb-patterning: candidate pathways for the origin and evolutionary diversification of beetle ‘horns’. Heredity 97: 179–191.
11. KotiahoJS (2000) Testing the assumptions of conditional handicap theory: costs and condition dependence of a sexually selected trait. Behav Ecol Sociobiol 48: 188–194.
12. BondurianskyR, RoweL (2005) Intralocus sexual conflict and the genetic architecture of sexually dimorphic traits in Prochyliza xanthostoma (Diptera: Piophilidae). Evolution 59: 1965–1975.
13. KoppA (2012) Dmrt genes in the development and evolution of sexual dimorphism. Trends Genet 28: 175–184.
14. MatsonCK, ZarkowerD (2012) Sex and the singular DM domain: insights into sexual regulation, evolution and plasticity. Nat Rev Genet 13: 163–174.
15. ClineTW, MeyerBJ (1996) Vive la difference: males vs females in flies vs worms. Annu Rev Genet 30: 637–702.
16. OhbayashiF, SuzukiMG, MitaK, OkanoK, ShimadaT (2001) A homologue of the Drosophila doublesex gene is transcribed into sex-specific mRNA isoforms in the silkworm, Bombyx mori. Comp Biochem Physiol B Biochem Mol Biol 128: 145–158.
17. SuzukiMG, FunagumaS, KandaT, TamuraT, ShimadaT (2005) Role of the male BmDSX protein in the sexual differentiation of Bombyx mori. Evol Dev 7: 58–68.
18. HasselmannM, GempeT, SchiøttM, Nunes-SilvaCG, OtteM, et al. (2008) Evidence for the evolutionary nascence of a novel sex determination pathway in honeybees. Nature 454: 519–522.
19. HedigerM, HenggelerC, MeierN, PerezR, SacconeG, et al. (2010) Molecular characterization of the key switch F provides a basis for understanding the rapid divergence of the sex-determining pathway in the housefly. Genetics 184: 155–170.
20. KijimotoT, MoczekAP, AndrewsJ (2012) Diversification of doublesex function underlies morph-, sex-, and species-specific development of beetle horns. Proc Natl Acad Sci U S A 109: 20526–20531.
21. ShuklaJN, PalliSR (2012) Doublesex target genes in the red flour beetle, Tribolium castaneum. Sci Rep 2: 00948 Available: http://www.nature.com/srep/2012/121210/srep00948/full/srep00948.html.
22. ItoY, HarigaiA, NakataM, HosoyaT, ArayaK, et al. (2013) The role of doublesex in the evolution of exaggerated horns in the Japanese rhinoceros beetle. EMBO Rep 14: 561–567.
23. ErdmanSE, ChenHJ, BurtisKC (1996) Functional and genetic characterization of the oligomerization and DNA binding properties of the Drosophila doublesex proteins. Genetics 144: 1639–1652.
24. BakerBS, WolfnerMF (1988) A molecular analysis of doublesex, a bifunctional gene that controls both male and female sexual differentiation in Drosophila melanogaster. Genes Dev 2: 477–489.
25. BurtisKC, BakerBS (1989) Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell 56: 997–1010.
26. ValenaS, MoczekAP (2012) Epigenetic mechanisms underlying developmental plasticity in horned beetles. Genet Res Int 2012 Article ID 576303. Available: http://dx.doi.org/10.1155/2012/576303.
27. Darwin C (1871) The descent of man and selection in relation to sex. London: John Murray.
28. HuxleyJS (1931) Relative growth of mandibles in stag beetles (Lucanidae). J Linn Soc Lond 37: 675–703.
29. HosoyaT, ArayaK (2005) Phylogeny of Japanese stag beetle (Coleoptera: Lucanidae) inferred from 16S mtrRNA gene sequences, with reference to the evolution of sexual dimorphism of mandibles. Zool Sci 22: 1305–1318.
30. KawanoK (2006) Sexual dimorphism and the making of oversized male characters in beetles (Coleoptera). Ann Entomol Soc Am 99: 327–341.
31. Kodric-BrownA, SiblyRM, BrownJH (2006) The allometry of ornaments and weapons. Proc Natl Acad Sci U S A 103: 8733–8738.
32. Fujita H (2010) The Lucanid Beetles of the world. Tokyo: Mushi-sha. (In Japanese).
33. GotohH, FukayaK, MiuraT (2012) Heritability of male mandible length in the stag beetle Cyclommatus metallifer. Entomol Sci 15: 430–433.
34. OliveiraDCSG, WerrenJH, VerhulstEC, GiebelJD, KampingA, et al. (2009) Identification and characterization of the doublesex gene on Nasonia. Insect Mol Biol 18: 315–324.
35. BayrerJR, ZhangW, WeissMA (2005) Dimerization of doublesex is mediated by a cryptic ubiquitin-associated domain fold: implications for sex-specific gene regulation. J Biol Chem 280: 32989–32996.
36. RobinettCC, VaughanAG, KnappJM, BakerBS (2010) Sex and the single cell. II. There is a time and place for sex. PLoS Biol 8(5): e1000365 Available: http://www.plosbiology.org/article/info%3Adoi%2F10.1371%2Fjournal.pbio.1000365.
37. TanakaK, BarminaO, SandersLE, ArbeitmanMN, KoppA (2011) Evolution of sex-specific traits through changes in HOX-Dependent doublesex expression. PLoS Biol 9(8): e1001131 Available: http://www.plosbiology.org/article/info%3Adoi%2F10.1371%2Fjournal.pbio.1001131.
38. Nijhout HF (1994) Insect hormones. Princeton: Princeton Univ. Press.
39. EmlenDJ, NijhoutHF (1999) Hormonal control of male horn length dimorphism in the dung beetle Onthophagus Taurus (Coleoptera: Scarabaeidae). J Insect Physiol 45: 45–53.
40. FryCL (2006) Juvenile hormone mediates a trade-off between primary and secondary sexual traits in stalk-eyed flies. Evol Dev 8: 191–201.
41. NiitsuS, LobbiaS, KamitoT (2011) In vitro effects of juvenile hormone analog on wing disc morphogenesis under ecdysteroid treatment in the female-wingless bagworm moth Eumeta variegate (Insecta: Lepidoptera, Psychidae). Tissue Cell 43: 143–150.
42. ShingletonAW, DasJ, ViniciusL, SternDL (2005) The temporal requirements for insulin signaling during development in Drosophila. PLoS Biol 3(9): e289 Available: http://www.plosbiology.org/article/info%3Adoi%2F10.1371%2Fjournal.pbio.0030289.
43. EdgarBA (2006) How flies get their size: Genetics meets physiology. Nat Rev Genet 7: 907–916.
44. TangHY, Smith-CaldasMSB, DriscollMV, SalhadarS, ShingletonAW (2011) FOXO regulates organ-specific phenotypic plasticity in Drosophila. PLoS Genet 7(11): e1002373 Available: http://www.plosgenetics.org/article/info%3Adoi%2F10.1371%2Fjournal.pgen.1002373.
45. LarkinMA, BlackshieldsG, BrownNP, ChennaR, McGettiganPA, et al. (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23: 2947–2948.
46. R Core Team (2012) R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. Available: http://R-project.org/.
47. BenjaminiY, HochbergY (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol 57: 289–300.
Štítky
Genetika Reprodukčná medicínaČlánok vyšiel v časopise
PLOS Genetics
2014 Číslo 1
- Je „freeze-all“ pro všechny? Odborníci na fertilitu diskutovali na virtuálním summitu
- Gynekologové a odborníci na reprodukční medicínu se sejdou na prvním virtuálním summitu
Najčítanejšie v tomto čísle
- GATA6 Is a Crucial Regulator of Shh in the Limb Bud
- Large Inverted Duplications in the Human Genome Form via a Fold-Back Mechanism
- Differential Effects of Collagen Prolyl 3-Hydroxylation on Skeletal Tissues
- Affects Plant Architecture by Regulating Local Auxin Biosynthesis