
The Cohesin Subunit Rad21 Is Required for Synaptonemal Complex Maintenance, but Not Sister Chromatid Cohesion, during Drosophila Female Meiosis
Meiosis is a specialized form of cell division that ensures production of germ cells with the right number of chromosomes, so that at fertilization the embryo receives complete sets of paternal and maternal chromosomes. The accurate distribution of chromosomes during cell divisions is dependent on a ring-shaped protein complex called cohesin. Cohesin is thought to embrace the chromosomes from the time of their duplication during S-phase until their segregation in the ensuing division. This segregation is facilitated by the controlled proteolytic cleavage of one of the cohesin ring components. Most eukaryotes express specialized variants of this protein: for mitosis the variant Rad21/Scc1/Mcd1 and for meiosis the related protein Rec8. Because Drosophila lacks a clear Rec8 homolog, we have analyzed in the present study whether the mitotic variant Rad21 may also function during meiosis. We have destroyed Rad21-based cohesin by premature cleavage of an engineered Rad21 variant during oogenesis. While we find no indication for effects on the accuracy of meiotic chromosome segregation, Rad21 cleavage results in a premature disassembly of the synaptonemal complex (SC), a structure required for meiotic recombination in Drosophila oocytes. Our interaction studies provide intriguing hints how Rad21 might contribute to SC maintenance.
Published in the journal:
The Cohesin Subunit Rad21 Is Required for Synaptonemal Complex Maintenance, but Not Sister Chromatid Cohesion, during Drosophila Female Meiosis. PLoS Genet 10(8): e32767. doi:10.1371/journal.pgen.1004540
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1004540
Summary
Meiosis is a specialized form of cell division that ensures production of germ cells with the right number of chromosomes, so that at fertilization the embryo receives complete sets of paternal and maternal chromosomes. The accurate distribution of chromosomes during cell divisions is dependent on a ring-shaped protein complex called cohesin. Cohesin is thought to embrace the chromosomes from the time of their duplication during S-phase until their segregation in the ensuing division. This segregation is facilitated by the controlled proteolytic cleavage of one of the cohesin ring components. Most eukaryotes express specialized variants of this protein: for mitosis the variant Rad21/Scc1/Mcd1 and for meiosis the related protein Rec8. Because Drosophila lacks a clear Rec8 homolog, we have analyzed in the present study whether the mitotic variant Rad21 may also function during meiosis. We have destroyed Rad21-based cohesin by premature cleavage of an engineered Rad21 variant during oogenesis. While we find no indication for effects on the accuracy of meiotic chromosome segregation, Rad21 cleavage results in a premature disassembly of the synaptonemal complex (SC), a structure required for meiotic recombination in Drosophila oocytes. Our interaction studies provide intriguing hints how Rad21 might contribute to SC maintenance.
Introduction
During meiosis, haploid germ cells are generated from diploid parental cells by two consecutive cell divisions without intervening DNA replication. Before the first meiotic division, homologous chromosomes are paired into bivalents and the two sister centromeres in each homolog are constrained to behave as a functional unit. The two homologous centromeres of each bivalent are bi-oriented in the spindle and segregated apart during the first meiotic division. Thereafter sister centromeres become functionally independent, allowing their bi-orientation and separation during the second meiotic division, very much like during mitosis (for review see: [1]). Importantly, error-free chromosome segregation during each meiotic division (homologs in meiosis I and sisters in meiosis II) does not just depend on regulated centromere behavior but also on temporal and regional control of sister chromatid cohesion.
Sister chromatid cohesion in combination with meiotic crossovers keeps bivalents physically together until the metaphase-to-anaphase transition of the first meiotic division. Crossovers are generated by meiotic recombination between non-sister chromatids of homologous chromosomes. The order of events during initiation of meiotic recombination varies among the organisms. In mice, fungi and plants double strand breaks (DSBs) mark the first event of meiotic recombination, and DSBs are required for the intimate pairing (synapsis) of homologous chromosomes during the extended prophase of meiosis I. In Drosophila, however, synapsis can occur in the absence of prior DSB formation [2]. A unique proteinaceous structure, the synaptonemal complex (SC), is formed during early stages of prophase I between the homologs. SC formation commences with the establishment of the axial elements (AE) which represent a scaffold running alongside the paired sister chromatids within each homolog. Concomitant with pairing of homologs, the AE mature into the lateral elements (LE) of the SC. The LE are connected by perpendicularly oriented transverse filaments (TF) which form the central element (CE) of the SC. In Drosophila melanogaster, meiotic recombination only occurs in females and consequently the SC is only assembled during oogenesis. The protein C(2)M has been identified as a component of the LE, and the main element of the TF is the elongated coiled-coil protein C(3)G [3], [4]. Loss of either protein results in a severely compromised SC structure and high levels of chromosome non-disjunction during the meiotic divisions [3], [4]. Proper C(3)G localization requires C(2)M but not vice versa [3].
After crossover formation, the SC is disassembled and crossovers mature into chiasmata. Despite SC disassembly, paired homologous chromosomes cannot disjoin, because sister chromatid cohesion distal to the crossover sites prevents terminalization of chiasmata. This cohesion between replicated sister chromatids is mediated by the heterotetrameric ring-shaped cohesin complex (for review see: [5], [6]). Cohesin complex components were originally identified by genetic screens in the yeast Saccharomyces cerevisiae [7], [8]. The core cohesin complex consists of the two structural maintenance of chromosomes (SMC) molecules SMC1 and SMC3, which form extended intramolecular coiled-coils and heterodimerize via their hinge regions. An α-kleisin subunit connects SMC1 and SMC3 by binding to their head domains, thus forming a tripartite ring-like structure. The α-kleisin Rad21/Scc1/Mcd1 also recruits the accessory subunit Scc3 (for reviews see: [5], [6]). The cohesin ring most likely embraces the sister chromatids and thereby establishes sister chromatid cohesion topologically [9].
Several eukaryotes are known to express meiosis-specific cohesin components (for review see: [10]). In yeasts the meiosis-specific α-kleisin Rec8 associates with the SMC head domains instead of Rad21/Scc1/Mcd1 [11], [12]. Apart from Rec8 homologs, vertebrate genomes encode yet an additional meiosis-specific α-kleisin, Rad21L, but a direct role for this variant in sister chromatid cohesion awaits demonstration [13], [14], [15]. However, it has been shown in mouse spermatocytes that Rad21L is involved in assembly of the axial elements of the SC [16], [17]. An involvement of cohesin in SC maintenance has been demonstrated previously in several distantly related eukaryotes [11], [18], [19], [20], [21]. Mammalian meiotic cohesin complexes contain the specific subunit SMC1β and the Scc3 homolog STAG3/SA3, while mitotic cells harbor cohesin complexes with SMC1α and either STAG1/SA1 or STAG2/SA2. Not all imaginable combinations of these subunits may be realized in cohesin complexes occurring in vivo, but immunoprecipitation of complexes present in mouse testis extracts revealed five variant cohesin complexes with differing subunit composition [15].
Cohesion is abrogated at the metaphase-to-anaphase transition by proteolytic cleavage of the α-kleisin cohesin subunit by the cysteine protease separase, thus opening the cohesin ring and liberating the sister chromatids [22], [23]. In meiosis, two waves of separase activity occur during the two divisions. In meiosis I, separase-dependent cleavage of phosphorylated Rec8, which is present in cohesin complexes located at the chromosome arm regions, allows chiasmata terminalization and hence homolog separation [24], [25], [26], [27]. Importantly, Rec8 in cohesin complexes located within pericentromeric regions is protected from proteolytic cleavage during meiosis I. Proteins of the Shugoshin (Sgo)-family recruit protein phosphatase 2A (PP2A) to the centromeric region, thus keeping Rec8 locally in a cleavage-resistant unphosphorylated state [28], [29]. Consequently, sister centromeres remain paired throughout meiosis I permitting their bi-orientation during meiosis II. A second burst of separase activity destroys pericentromeric cohesion before anaphase II.
In several organisms, the α-kleisin Rad21 is expressed not only before mitotic but also before meiotic divisions. Its role in meiotic sister chromatid cohesion has been discussed controversially. In the mouse, Rad21 is clearly expressed in meiotic cells of both sexes [30]. A number of immunolocalization studies have shown the persistence of Rad21 on mammalian meiotic chromatin at least through meiosis I which has been interpreted as Rad21 possibly serving a cohesive function during meiosis [20], [31], [32], [33], [34]. However, other studies either failed to detect Rad21 in premeiotic S-phase or later stages of rat spermatogenesis [35], or reported Rad21 to apparently leave chromatin before metaphase I in mouse spermatocytes [36]. Elegant functional studies have recently revealed that premature TEV protease-mediated cleavage of all Rad21 has no obvious effect on chromatid cohesion in mouse oocytes, while analogous premature Rec8 cleavage resulted in premature and complete loss of cohesion both in metaphase I, leading to chiasmata resolution, and also in metaphase II [37]. Thus, Rad21 serves no cohesive function during meiosis, at least not in mouse oocytes, and Rec8 cleavage is sufficient for loss of cohesion in both meiotic divisions.
Intriguingly, the Drosophila genome does not contain an obvious Rec8 homolog. However, the SC component C(2)M [3] was shown to be a divergent member of the α-kleisin family by in-depth bioinformatics analyses [38]. Its meiosis-specific expression, its association with the cohesion subunit SMC3, and the high level of chromosome missegregation in c(2)M mutants are consistent with C(2)M functioning analogous to Rec8. However, the low level of sister non-disjunction in c(2)M mutants, as well as C(2)M localization dynamics during meiosis and the lack of abnormalities after expression of variants predicted to be separase cleavage-resistant, argue against C(2)M being the bona fide Rec8 homolog [3], [39]. Two additional genes that seem to be specific to the Drosophila lineage, solo and ord, qualify to encode meiotic cohesins, as both solo and ord null mutants show premature dissociation of homologous chromosomes and of sister chromatids, resulting in high frequencies of meiotic non-disjunction events [40], [41], [42]. Also, in ord and solo mutants, the SC is formed, but it disassembles prematurely [41], [43]. However, neither SOLO nor ORD display similarity with α-kleisins at the primary structure level and there are no reports that either one of the two proteins is a substrate for separase, which is active during the meiotic divisions in Drosophila [39]. Thus, it is at present not clear whether Drosophila harbors, as part of meiotic cohesin complexes, an α-kleisin homolog, which needs to be removed in a separase-dependent manner during the meiotic divisions to allow chromosome/chromatid segregation.
Here we have addressed whether Drosophila Rad21/Verthandi takes over the function of Rec8 by acting as a meiotic α-kleisin and whether it is involved in SC maintenance. We find that experimentally induced premature Rad21 proteolysis during oogenesis does not result in premature chromosome/chromatid separation or chromosome non-disjunction, arguing against an essential contribution of Rad21 to meiotic sister chromatid cohesion. However, maintenance of the SC is clearly dependent on Rad21, which co-localizes with C(2)M and C(3)G in nuclei with a fully formed SC. Moreover, our finding that C(2)M can interact physically with Rad21 allows speculations towards a molecular mechanism for the linkage of the SC to meiotic chromosome cores in Drosophila.
Results
Targeted Rad21 inactivation during female meiosis
Rad21 provides essential functions during mitosis. To evaluate whether Rad21 also provides important functions during meiosis, we applied a system allowing controlled Rad21 inactivation specifically during oogenesis. We took advantage of Drosophila strains expressing Rad21 variants that can be proteolytically inactivated by TEV protease. These Rad21 variants contain three consecutive TEV protease cleavage sites at position 271 or 550, as well as a C-terminal myc-epitope tag (Rad21TEV-myc). Rad21 mutant rescue experiments have proven these variants to be functional [44]. Furthermore, TEV protease expression has been shown to result in efficient Rad21TEV-myc cleavage and consequential inactivation. Cleavage in mutant embryos that rely on Rad21TEV-myc as their sole Rad21 species, resulted in completely penetrant premature sister chromatid separation during the first mitosis after onset of TEV protease expression [44].
To express specifically in the female germline a UAS transgene, which encodes a TEV protease variant with improved catalytic properties (see materials and methods), we used the maternal alpha-tubulin GAL4-driver line (mat-GAL4). The resulting efficiency of Rad21TEV-myc cleavage was assessed with extracts from stage 14 oocytes. Oocytes from sibling females with and without UAS-TEV protease transgene were compared (Fig. 1A, +TEV and −TEV). Immunoblot analyses using antibodies against myc allowed the detection of full-length Rad21TEV-myc as well as the C-terminal cleavage product (Fig. 1B). Quantification revealed that around 95% of Rad21TEV-myc was cleaved in the TEV protease expressing oocytes (Fig. 1B).
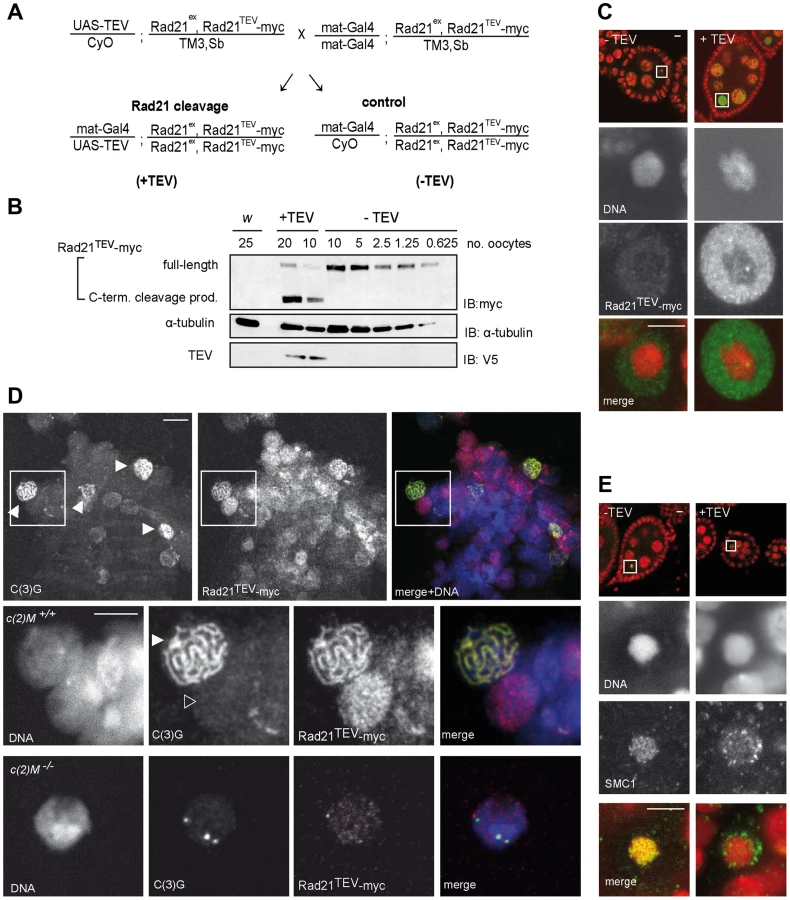
To assess phenotypic consequences of Rad21TEV-myc cleavage in situ, we immunolabeled ovarioles with anti-myc antibodies. In the absence of TEV protease expression, only weak signals were obtained within the nucleoplasm surrounding the karyosome, the highly condensed chromatin of the oocytes (Fig. 1C, −TEV). However, upon expression of TEV protease, strong anti-myc signals were detected in the nucleoplasm (Fig. 1C, +TEV), indicative of Rad21TEV-myc cleavage product accumulation. The inability to detect uncleaved Rad21TEV-myc localizing on oocyte chromatin in these whole mount preparations could, in principle, be due to accessibility problems. Thus, we also stained chromosome spread preparations of germaria and early egg chambers. While this method did not allow the unambiguous assignment of nuclei to specific stages of oogenesis, we clearly detected nuclear anti-myc signals in cells of germaria. Most importantly, while the signals were diffuse in nuclei without an SC, we obtained strong anti-myc signals co-localizing with the synaptonemal complex (SC) component C(3)G in the typical thread-like pattern in pro-oocytes (Fig. 1D). To confirm the presence of Rad21 in the SC, we also analyzed spread preparations of ovarioles from females expressing a functional Rad21-EGFP variant by double labeling with anti-EGFP and anti-C(3)G antibodies. We again observed co-localization in nuclei with a fully formed SC, corroborating our results obtained for localization of Rad21TEV-myc.
Rad21 is, together with SMC1 and SMC3, part of the tripartite cohesin ring, embracing sister chromatids after DNA replication (for review see [5]). As the cohesin rings are bound to chromatin in a topological fashion, SMC1 and SMC3 are expected to dissociate from chromatin upon Rad21TEV-myc cleavage. In contrast to Rad21, SMC1 and SMC3 can be readily visualized on meiotic chromatin by immunostaining of Drosophila ovariole whole mount preparations [45]. There, SMC1 and SMC3 are associated with the lateral elements of the SC [45]. Indeed, while a characteristic pattern of SMC1 can be detected in the karyosome in the absence of TEV protease expression, SMC1 is delocalized upon Rad21TEV-myc cleavage (Fig. 1E). We conclude that in our system Rad21TEV-myc is efficiently cleaved during oogenesis, and that this cleavage leads to premature dissociation of cohesin from meiotic chromatin.
Premature Rad21 cleavage causes precocious disassembly of the synaptonemal complex
Since Rad21TEV-myc cleavage occurs during a developmental stage when the SC is fully formed in the oocyte nucleus (TEV protease expression driven by mat-GAL4 can be detected starting in region 3 of the germaria; Fig. S1A), we addressed possible phenotypic consequences on SC integrity. Immunolabeling of the SC-components C(3)G and an HA tagged variant of C(2)M within wild type oocyte nuclei of stage 4–5 egg chambers resulted in the expected ribbon-like SC staining (Fig. 2) [3], [4]. TEV protease expression in a background without Rad21TEV-myc but with wild type Rad21 did not affect the SC-associated anti-C(3)G signals that were just like in wild type ovarioles (Fig. 2A). A normal C(3)G staining pattern was also observed in control ovarioles expressing Rad21TEV-myc in the Rad21 mutant background in the absence of TEV protease (Fig. 2A), and in ovarioles from Rad21+/Rad21− heterozygote individuals (Fig. 3B). However, the characteristic ribbon-like C(3)G staining was almost completely lost from the oocyte chromatin after TEV protease expression in a background with exclusively Rad21TEV-myc, and C(3)G accumulated instead in the nucleoplasm (Fig. 2A). We point out that in these same ovarioles at earlier stages within the germarium, C(3)G staining still revealed the normal ribbon-like structures (Fig. S2), as expected, because mat-GAL4 driven TEV protease expression is not yet detectable at these early stages (Fig. S1A).
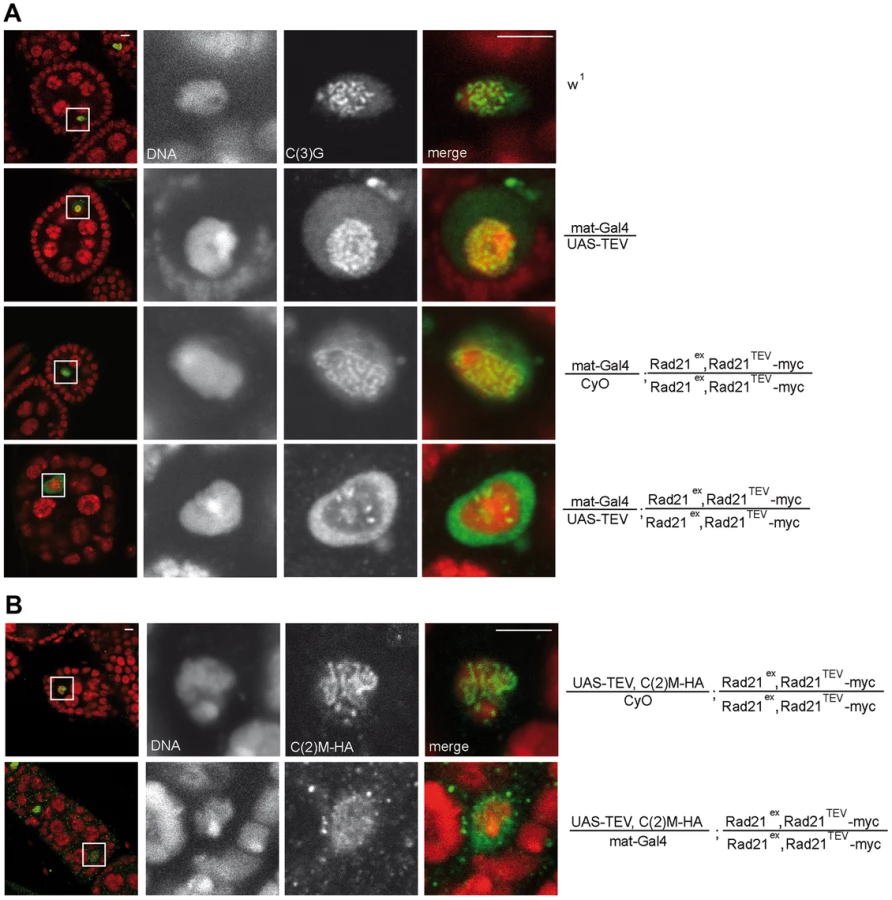

To assess whether the localization of the lateral SC component C(2)M is also affected after premature Rad21 cleavage, we generated flies, which express c(2)M-HA under genomic control in a rad21 mutant background rescued by Rad21TEV-myc expression. When TEV protease was expressed in these ovarioles, the ribbon-like C(2)M-HA-staining typical for the SC also disappeared from the meiotic chromatin and C(2)M-HA distributed throughout the nucleus (Fig. 2B).
Thus, our results suggest that the SC disassembles as a consequence of Rad21TEV-myc cleavage. To evaluate whether the observed phenotype is due to a dominant negative effect of the particular cleavage products generated, we analyzed the dependence of the phenotype on the precise position of the TEV cleavage sites within Rad21TEV-myc. Moreover, to rule out effects of the GAL4 driver background, we repeated the experiments with a different driver, nanos (nos)-GAL4. nos-GAL4 expression commences earlier during oogenesis, in region 2a of the germarium (Fig. S1B). Indeed, TEV protease expression directed by this driver resulted in premature SC disassembly at an even earlier stage during oogenesis (nos-GAL4: stage 3.4+/−0.6 (n = 34); mat-GAL4: stage 5.3+/−0.6 (n = 30); P<0.0001; Mann-Whitney U-test; Fig. 3A). Moreover, SC disassembly was observed to occur at an earlier stage with Rad21TEV-myc having the TEV cleavage sites after amino acid 271 compared to after amino acid 550 (position 271: SC disassembly + TEV at stage 3.4+/−0.6 vs. − TEV at stage 7.4+/−0.5; position 550: SC disassembly + TEV at stage 5.4+/−0.6 vs. −TEV stage 7.1+/−0.6; Fig. 3A).
Although nos-GAL4 driven TEV protease expression can be detected early in region 2a of the germarium, establishment of the SC was not affected (Fig. S2). Analysis of spread preparations of germaria revealed clear evidence of initial Rad21 degradation before SC disassembly (Fig. S3). Therefore, the SC disassembly which is observed during later oogenesis might depend on complete Rad21 degradation.
Taken together, our results demonstrate that precocious SC disassembly is a robust phenotype that is observed with different GAL4 drivers and different TEV cleavage site insertion positions within Rad21TEV-myc.
Interestingly, premature SMC1 delocalization and SC disassembly also occurred when Rad21TEV-myc was cleaved by TEV protease expression in the presence of one wild type Rad21+ allele (Fig. 3). While in these cases the SC stayed intact longer than in the Rad21 mutant situation, the difference in SC disassembly timing compared to the control situation was still highly significant (position 271: stage 5.4+/−0.8 vs. stage 7.4+/−0.5; position 550: stage 6.1+/−0.5 vs. stage 7.1+/−0.6; Fig. 3A). The precocious SC disassembly is not due to the reduced Rad21+ gene dosage, because the dynamics of SC disassembly is like wild type in females heterozygous for Rad21ex without any ectopic Rad21TEV-myc cleavage (Fig. 3). Moreover, we re-iterate that TEV protease expression in a background with wild type Rad21 has no effect when Rad21TEV-myc is not expressed (Fig. 2A and 3A).
As an independent approach to remove Rad21 from developing egg chambers, we applied targeted destruction of GFP-tagged proteins by the deGradFP system [46]. In this system, GFP-fused proteins are recruited to a recombinant SCF ubiquitin ligase generated by expression of a specific single-chain anti-GFP antibody fused to the F-box region of Slmb (NSlmb-vhhGFP4). The recruitment of GFP fusions by NSlmb-vhhGFP4 results in their proteasomal degradation. We constructed strains in which Rad21 mutants are rescued by the expression of Rad21-EGFP. NSlmb-vhhGFP4 expression driven by mat-GAL4 markedly reduced Rad21-EGFP protein levels (Fig. S4). nanos-GAL4 driven expression of the NSlmb-vhhGFP4 fusion protein again resulted in premature dissociation of the SC. The difference in SC disassembly timing compared to the control situation (Rad21ex, Rad21-EGFP homozygous females without NSlmb-vhhGFP4 expression) was again highly significant (Rad21ex, Rad21-EGFP homozygous females + NSlmb-vhhGFP4: stage 5.6+/−0.8 vs. stage 7.2+/−0.5; Rad21ex, Rad21-EGFP heterozygous females + NSlmb-vhhGFP4: stage 6.5+/−0.5 vs. stage 7.1+/−0.5; Fig. 3A). This premature SC disassembly after proteasomal degradation of Rad21-EGFP confirms that loss of Rad21 results in SC disintegration. This SC instability therefore does not depend on the presence of Rad21TEV-myc cleavage fragments, which are generated after TEV protease expression, and which might in principle have a dominant effect.
Although the majority of the SC components C(3)G and C(2)M leaves the oocyte chromatin after forced Rad21 cleavage, some bright staining patches remain (Fig. 2A, Fig. 3A). As residual SC components have been described to remain associated with clustered centromeres after normal SC disassembly [47], we analyzed whether residual C(3)G after ectopic Rad21TEV-myc cleavage was colocalized with centromeres. Upon premature Rad21 cleavage, we indeed found colocalization of the centromeric H3 variant Cid/Cenp-A with persisting C(3)G patches (Fig. S5), suggesting that association of the SC with centromeric regions might not depend on Rad21-containing cohesin. Taken together, our data imply that the integrity of the mitotic α-kleisin cohesin subunit Rad21 is required for SC maintenance at chromosome arms during Drosophila oogenesis.
Rad21 binds to the N terminus of the SC component C(2)M
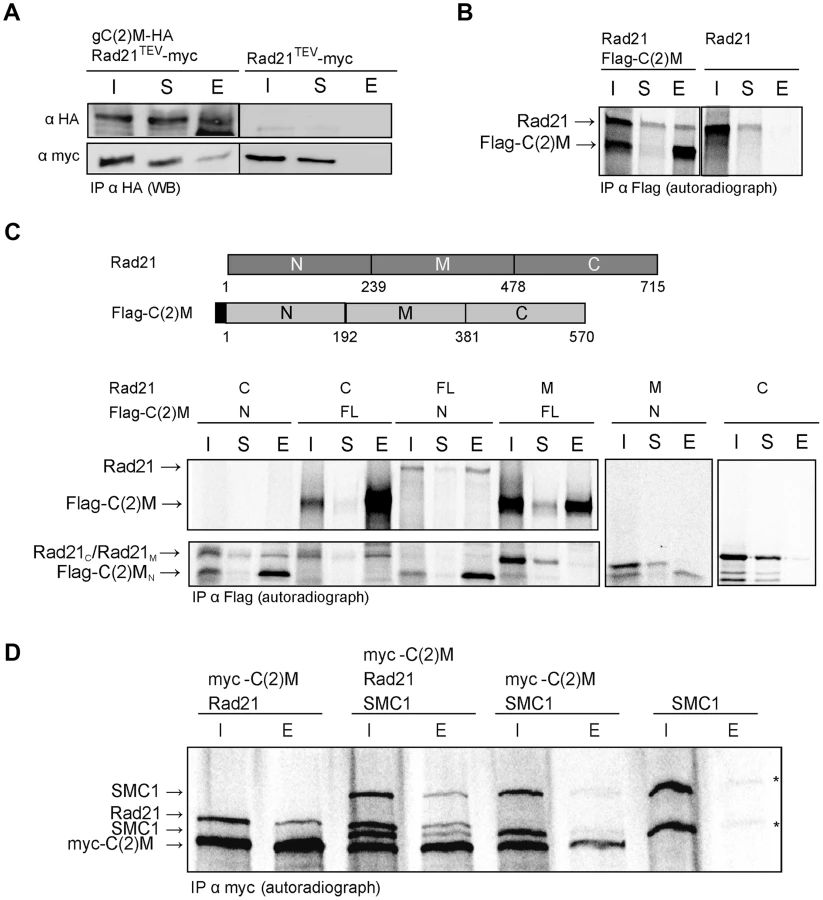
To address how Rad21 interacts with the SC, we first analyzed whether Rad21 protein might bind to the SC component C(2)M. We performed co-immunoprecipitation experiments using protein extracts prepared from fly embryos expressing c(2)M-HA and Rad21TEV-myc. The presence of C(2)M and Rad21 in early embryos has been demonstrated previously [39]. Indeed, Rad21TEV-myc was co-precipitated with C(2)M-HA. In control experiments, where we used the same anti-HA antibodies for immunoprecipitation from an extract containing Rad21TEV-myc, but not C(2)M-HA, we were unable to pull down Rad21TEV-myc (Fig. 4A), ruling out a non-specific association of Rad21TEV-myc with HA-antibodies or beads. To obtain independent support for an interaction between Rad21 und C(2)M, we conducted in vitro pull-down assays. To this end, we used an in vitro transcription/translation (IVT) system to synthesize Rad21 and Flag epitope-tagged C(2)M in a reticulocyte lysate in the presence of [35S]methionine. Autoradiography of the samples after anti-Flag immunoprecipitation revealed that Rad21 specifically bound to Flag-C(2)M (Fig. 4B). To delineate the interacting domains of the two proteins, we repeated the assay with in vitro synthesized fragments of both proteins. These experiments revealed that an N-terminal fragment of C(2)M (C(2)MN, aa 1–191) is sufficient to precipitate Rad21 (Fig. 4C). Furthermore, the C-terminus of Rad21 (Rad21C, aa 478–715) is sufficient for interaction with Flag-C(2)M or Flag-C(2)MN (Fig. 4C). None of the other fragments were able to mediate an interaction in this system (Fig. 4C). In the same assay system, we neither detected an interaction between Rad21 and one of the other SC components, C(3)G or Corona, nor an interaction between Rad21 and the cohesion proteins ORD or SOLO (data not shown). C(2)M has been found in a complex with SMC3 [39]. If C(2)M binds directly to the SMC heads, as it is regarded typical for α-kleisins, one would expect the binding of C(2)M and Rad21 to the SMC cohesin subunits to be mutually exclusive. Using the IVT system, we analyzed the binding potential of C(2)M and Rad21 towards SMC1. In these experiments, C(2)M was N-terminally fused with a 6× myc epitope tag, co-expressed with Rad21 and/or SMC1 and anti-myc immunoprecipitates were analyzed (Fig. 4D). While Rad21 could be readily co-immunoprecipitated together with myc-C(2)M, co-immunoprecipitation of SMC1 depended on the presence of Rad21. Thus, these data suggest that myc-C(2)M does not bind directly to SMC1 but that Rad21 mediates the association of C(2)M with SMC1. Taken together, our immunoprecipitation analyses reveal a novel interaction between the two α-kleisin proteins Rad21 and C(2)M. Specifically, we show that the C-terminus of Rad21 binds to the N-terminus of C(2)M, suggesting that this interaction mediates the association of C(2)M with the core cohesin complex. In vivo, localization of Rad21 and C(2)M are mutually dependent, consistent with an interaction of these two proteins (Figs. 1D and 2B).
Rad21 cleavage does not profoundly affect cohesion during the meiotic divisions
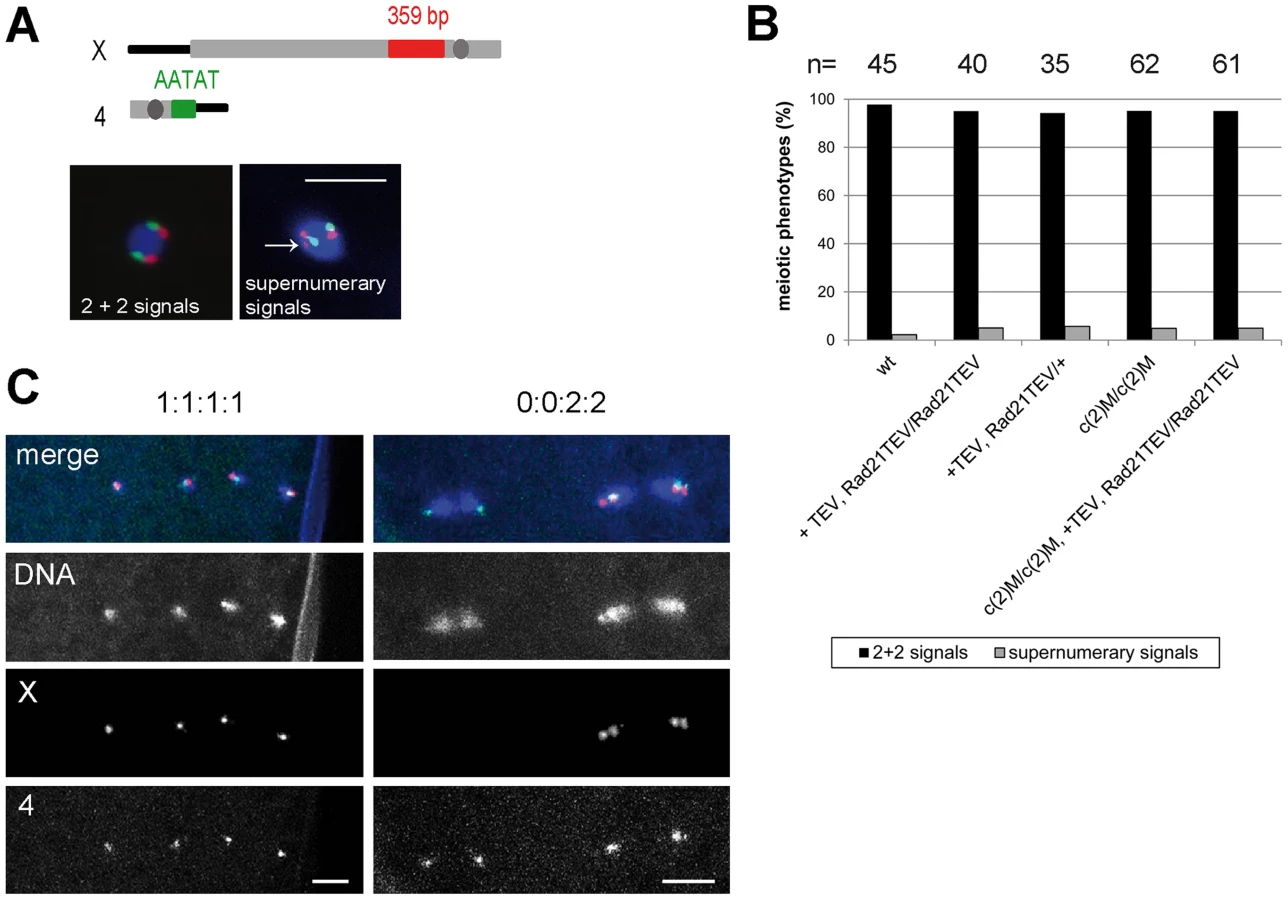
Having established that Rad21TEV-myc cleavage results in premature SC disassembly, we wondered whether additional late meiotic processes were affected. If Rad21 is required for cohesion between sister chromatids during the meiotic divisions, one would expect precocious separation of sister chromatids in the Rad21 mutant situation, and consequently chromosome missegregation. Classical genetic non-disjunction assays are not possible in our system, because TEV protease expression in our experiments inactivates the essential maternal Rad21 contribution and therefore results in complete female sterility. In multiple experiments, after mat-GAL4 driven TEV protease expression causing Rad21TEV-myc cleavage, no larvae hatched from the eggs laid by those females. Immunofluorescence analysis of these embryos revealed massive defects already during the very early zygotic divisions. Fragmented and unequally sized DNA masses could be observed, organizing multiple and/or multipolar spindles (Fig. S6). Most embryos appeared to have arrested in a metaphase-like state. Thus, the sterility of these females precluded scoring of genetic markers in adult progeny. As an alternative approach, we applied fluorescent in situ hybridization (FISH) to detect chromosome-specific regions in metaphase I-arrested oocytes and analyzed them with reference to precocious chromosome separation. We used an X-chromosome-specific probe (359 bp repeat) and a chromosome 4-specific probe (AATAT)6 (Fig. 5A). The observed phenotypes were assigned to two different categories: (1) normal meiotic figures exhibiting 2+2 FISH signals, and (2) precocious separation of chromatids as indicated by supernumerary FISH signals (>2 FISH signals for at least one of the probes; Fig. 5A). In the wild type situation, 98% of the oocytes showed normal meiotic figures, as indicated by the two signals for the different chromosomes (Fig. 5A and B). 2% of the wild type oocytes contained more than 2 signals for one of the two probes (n = 45). After TEV protease-mediated Rad21TEV-myc cleavage, the distribution of phenotypes was similar: 95% of the oocytes displayed a normal arrangement, and 5% of the analyzed oocytes had an elevated number of FISH signals indicative of premature chromatid separation (n = 40). Also, oocytes from c(2)M mutant females which have been shown previously to display high levels of meiosis I non-disjunction [3], [39], showed no increase of premature chromatid separation (5% of the c(2)M mutant oocytes exhibited supernumerary FISH signals (n = 62). Finally, we analyzed oocytes of individuals in which Rad21TEV-myc cleavage was performed in the c(2)M mutant background (Fig. 5B). In this constellation, again, only 5% of the oocytes were assigned to the ‘supernumerary FISH signals’ category (n = 61). On the contrary, very similar analyses in ord mutants using a probe directed against the same repetitive region of the X-chromosome revealed a high proportion (46%) of prometaphase oocyte nuclei with three or four FISH signals, indicating loss of cohesion [48]. Thus, our data do not suggest any additional contribution of Rad21 to chromatid cohesion at this developmental stage during oogenesis. To analyze meiotic divisions directly, we also performed FISH on very early embryos shortly after egg deposition. In this experiment, we observed in the majority of cases a correct 1∶1∶1∶1 distribution of FISH signals among the four meiosis II products in the Rad21 mutant situation, indicative of normal segregation in both meiotic divisions. In only one out of 83 cases, we detected a clear example of missegregation (in meiosis I) with a signal distribution of 0∶0∶2∶2 (Fig. 5C). Taken together, after efficient cleavage of Rad21TEV-myc in the oocytes, the effects on meiotic chromosome segregation, if any, were very mild. These findings indicate that Rad21 is not required for sister chromatid cohesion in the oocyte nuclei, in contrast to ORD and SOLO [40], [41].
ORD and/or SOLO may function to maintain sister chromatid cohesion and thereby explain that Rad21 is not required during the meiotic divisions for normal chromosome segregation. To evaluate this possibility, we analyzed the localization of a functional Venus-SOLO variant [42]. In wild-type egg chambers, Venus-SOLO is localized in the vicinity of centromere clusters [41]. Upon Rad21TEV-myc cleavage, Venus-SOLO persisted in a dot-like pattern co-localizing with Cid/Cenp-A and C(3)G remnants (Fig. S7). In contrast, the ribbon-like C(3)G staining characteristically present in early egg chambers in wild type [4] (Fig. 2) was largely dissipated in the oocyte nucleoplasm, confirming that premature SC disassembly after Rad21TEV-myc cleavage occurred also in the Venus-SOLO background as expected (Fig. S7). The observed pericentromeric presence of SOLO even after Rad21TEV-myc cleavage is consistent with the notion that SOLO might render Rad21 dispensable during the meiotic divisions.
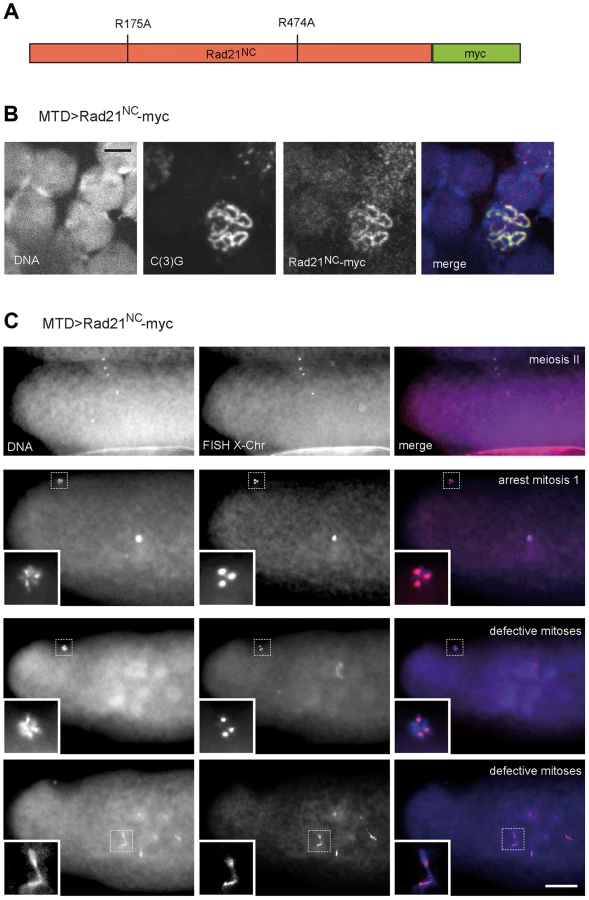
Alternatively, the apparently normal meiotic chromosome segregation observed after TEV protease-mediated Rad21TEV-myc cleavage before the meiotic divisions might also reflect the presence of a low, but sufficient, amount of residual non-cleaved Rad21TEV-myc. If sister chromatid cohesion during the meiotic division was indeed provided by Rad21 containing cohesin, separase-mediated Rad21 cleavage would be predicted to be essential for normal chromosome segregation during meiosis. To evaluate the significance of separase-mediated Rad21 cleavage during meiosis, we expressed a variant of Rad21-myc, in which the predicted separase cleavage sites (EXXR at positions 172–175 and 471–474) were destroyed by exchange of the arginines with alanines (Fig. 6A). This variant, dubbed Rad21NC-myc (non-cleavable), is predicted to be highly toxic in mitotically proliferating cells. Indeed, after expression of Rad21NC-myc in the proliferating eye imaginal disc, adults with severely reduced eyes were obtained (Fig. S8). After expression during oogenesis, Rad21NC-myc was observed to be co-localized with C(3)G in the SC during the early stages (Fig. 6B), indicating that this mutant is still capable to associate with chromatin. However, those females expressing Rad21NC-myc during oogenesis were almost completely sterile. Importantly, abnormalities were only apparent after normal completion of meiosis. All the late meiotic figures observed in early embryonic progeny were normal (13 clear MII anaphase/telophase figures among 230 analyzed embryos; Fig. 6C). FISH analysis demonstrated that X-chromosome segregation during meiosis is not perturbed by Rad21NC-myc expression (Fig. 6C). Apart from late meiotic figures, also all of the remnants of the polar bodies displayed a normal morphology and the expected three X chromosome FISH signals. In contrast to the meiotic divisions, however, mitotic divisions during early embryogenesis were severely affected by the maternally expressed Rad21NC-myc. In many embryos, strong defects were apparent already during mitosis 1, as only a single DNA mass was observed in the interior of the embryos (Fig. 6C). During this as well as later mitoses, prominent anaphase bridges were detected and X chromosome FISH revealed chromosome stretching (Fig. 6C), as expected after expression of a Rad21 variant that can no longer be cleaved by separase to initiate a normal anaphase. The observed drastic effect of Rad21NC-myc on mitotic, but not meiotic, chromosome segregation further confirms that Rad21 is not functioning as an essential α-kleisin component of meiotic cohesin.
Discussion
Rad21/Scc1 has been established as the α-kleisin subunit of cohesin in mitotic cycles from yeast to man. Even though Rad21/Scc1 is expressed during meiosis, a cohesive role in the meiotic divisions has been ruled out for murine female meiosis [37]. In Saccharomyces cerevisiae, Scc1 levels decline sharply when cells enter meiosis, while Rec8 abundance increases dramatically [11]. scc1 mutants have mild meiotic phenotypes and separase-dependent Rec8 cleavage is required for meiotic chromosome segregation [11], [26]. Likewise in Schizosaccharomyces pombe, Rec8, but not Rad21, localizes throughout chromatin during prophase of meiosis I and Rec8 cleavage is required for both meiosis I and meiosis II chromosome segregation [12], [27]. Thus, the emerging view is that during entry into the meiotic program a switch occurs from Rad21 containing cohesin complexes to Rec8 containing cohesin complexes, which are responsible for establishing and maintaining sister chromatid cohesion throughout meiosis. This initially simple picture has become more complicated in vertebrates with the discovery of Rad21L, whose possible function in sister chromatid cohesion remains to be addressed [13], [14], [15], [17]. The situation is even more puzzling in Drosophila, because an unambiguous Rec8 homolog appears to be missing and unrelated proteins like ORD and SOLO with no obvious homology to α-kleisins functionally qualify as cohesion proteins. Originally, C(2)M was assigned as the Drosophila Rec8 homolog based on its meiotic expression profile and its membership in the α-kleisin protein family [38]. However, we have shown that C(2)M accumulates on chromatin only after completion of premeiotic S-phase, appears to dissociate long before pro-metaphase I, and that the mutation of putative separase cleavage sites had no effect on C(2)M function, which is inconsistent with a behavior expected for a meiotic cohesin component [39]. Moreover, c(2)M mutants display high levels of non-disjunction only in meiosis I, and not in meiosis II, and SMC1/SMC3 is able to localize to meiotic chromatin in the absence of C(2)M [3], [45]. Thus, it remains an open question whether Drosophila expresses a meiotic α-kleisin, which needs to be removed in a stepwise fashion during the two meiotic divisions. In the present study, we have investigated whether Rad21 might function also as a meiotic kleisin in Drosophila, in addition to its established role as mitotic cohesin subunit. As a precedent, a recent study has shown that the protist Tetrahymena thermophila uses only one α-kleisin both in mitosis and meiosis [49].
If Rad21 fulfilled a cohesive function during the meiotic divisions in Drosophila, one would expect to observe after ectopic Rad21 cleavage a dissociation of paired homologous chromosomes and premature sister chromatid separation during the extended pachytene stage of meiosis I and, in addition, missegregation of chromosomes in both meiotic divisions. We have performed immunostainings against the constitutive Drosophila kinetochore component Cenp-C and we did not notice an elevated number of Cenp-C spots in the oocyte nuclei of early egg chambers after Rad21TEV-myc cleavage, arguing against premature chromatid separation. This conclusion is also supported by our FISH results where Rad21TEV-myc cleavage was not observed to cause increased missegregation during the meiotic divisions. In contrast, increased numbers of centromere signals and increased missegregation were clearly detected in ord and solo mutants, which lack proteins specifically required for meiotic chromatid cohesion [40], [41], [48].
We consider the explanation that the normal meiotic chromosome segregation observed after TEV protease-mediated premature cleavage of Rad21TEV-myc might be due to putative residual non-cleaved Rad21TEV-myc to be highly unlikely. The same experimental strategy has proven to be extremely efficient in case of mitosis [44]. When Rad21TEV-myc is the sole Rad21 species in mitotically proliferating cells, ectopic Rad21TEV-myc cleavage results in a completely penetrant premature separation of sister chromatids in the first mitosis following TEV protease expression. It could be argued that a meiosis-specific factor might shield Rad21TEV-myc from TEV protease-mediated cleavage. However, in mouse oocytes, TEV protease-mediated inactivation of the meiotic α-kleisin was demonstrated to be efficient, arguing against a conserved shielding mechanism [37], [50]. In addition, we point out that TEV protease-mediated cleavage of Rad21TEV-myc before the meiotic division destroyed the maternal contribution of this mitotic α-kleisin so effectively that embryonic mitoses were completely defective.
Our hypothesis that Rad21 does not act as an essential meiotic α-kleisin during Drosophila female meiosis not only rests on the evidence obtained by TEV protease-mediated premature Rad21TEV-myc cleavage, but also on the unperturbed meiotic chromosome segregation observed after expression of Rad21NC-myc, in which the separase consensus cleavage sites were destroyed by site-directed mutations. An assay for the direct biochemical analysis of Rad21 cleavage by separase is still lacking in the Drosophila system. Therefore, it remains to be shown whether Rad21NC-myc is indeed resistant to separase-dependent proteolysis. However, the consequences resulting from expression of this variant in mitotically proliferating cells are perfectly consistent with the presence of separase-resistant cohesin rings that cannot be opened at the metaphase-to-anaphase transition in mitosis to liberate, and allow segregation of, the replicated sister chromatids. After expression during oogenesis, Rad21NC-myc is incorporated into meiotic chromatin and it is present in amounts sufficient to inhibit early embryonic mitoses. Thus, if separase-dependent removal of Rad21-containing cohesin was a crucial step during meiotic chromosome segregation, severe phenotypic consequences should not be restricted to early embryonic mitoses. Meiotic chromosome segregation would be predicted to be affected as well. However, we did not observe any meiotic abnormalities like chromosome bridges or missegregation of the X-chromosome in the FISH analyses.
Proper SC assembly depends on all three known meiotic α-kleisins in C. elegans and on both Rad21L and Rec8 in mouse spermatocytes [16], [21], [51]. Also in yeasts, which express only one meiotic α-kleisin, Rec8, SC integrity depends on Rec8 [11], [52]. However, no role for the mitotic α-kleisin Rad21/Scc1 in maintaining SC integrity has been found so far. Thus, our observation that the SC disassembles prematurely upon Rad21TEV-myc cleavage in Drosophila, demonstrates for the first time the dependence of SC maintenance on intact, Rad21-containing cohesin. However, this premature SC disassembly does not result in chromosome missegregation later in meiosis. We assume that the premature SC disassembly induced in our experiments occurs not early enough to interfere with crossover formation. A normal presence of chiasmata might therefore explain the absence of chromosome segregation defects. While our results clearly demonstrate that Rad21 is required for maintenance of the SC, we did not see an effect on establishment of the SC, not even when we used nos-GAL4 to drive TEV protease expression early in the germarium. We suspect that nos-driven inactivation of Rad21TEV-myc is not fast or complete enough to profoundly affect SC maintenance in germarial stages, and only after a certain lag period enough TEV protease has accumulated to cleave sufficient Rad21TEV-myc, which then triggers SC disassembly.
An intriguing result of our experiments is the premature disassembly of the SC even when Rad21TEV-myc is ectopically cleaved, or when Rad21-EGFP is degraded, in the presence of one Rad21 wild type allele. Because SC disassembly follows wild type kinetics in Rad21 heterozygous females in the absence of ectopic Rad21 inactivation, the observed early SC disassembly after ectopic inactivation is not due to reduced Rad21 gene dosage. Also, because cleavage at different positions and ectopic degradation of Rad21-EGFP resulted in premature SC disassembly, a dominant negative effect of the Rad21 fragments on SC structure is highly unlikely. One possible explanation can be based on a model that more than one cohesin ring is required at each linkage position to tether the SC to the chromosome cores. If just one out of two (or more) interconnected cohesin rings is opened by TEV protease action, or Rad21-EGFP proteolysis, linkage at this point would be abrogated despite the presence of uncleaved Rad21 in interconnecting rings. In support of this model, interaction studies between cohesin subunits led to the proposal of a “handcuff model” postulating interconnected cohesin rings [53].
The interaction between C(2)M and Rad21, which this work has revealed, suggests a model how the SC might be linked to cohesin within the chromosome cores. We propose that a direct interaction between the α-kleisin proteins C(2)M and Rad21 may provide a structural framework within the SC. We point out that so far we have been unable to confirm this interaction, which we have detected by co-immunoprecipitation from embryonic extracts and in vitro translation reactions, also by co-immunoprecipitation from ovary extracts, presumably because of technical difficulties (expression levels, insolubility of the SC associated proteins). While our demonstration of the C(2)M-Rad21 interaction is, to our knowledge, the first published report of an association of different α-kleisins, a homodimerization of human Rad21 has been demonstrated using yeast two-hybrid assays and immunoprecipitation experiments [53]. The reported localization of C(2)M as an LE component of the SC is also consistent with a direct connection to cohesin, which localizes to the chromosome cores [3], [45], [54]. Electron microscopy (EM) studies have mapped the N-terminus of C(2)M to the inner edge of the LEs [54]. According to our data, we would also expect the C-terminus of Rad21 to localize to this region of the SC. Staining of chromosome squash preparations indeed revealed a clear co-localization of Rad21TEV-myc and C(3)G (Fig. 1D), very similar to what has been observed previously for SMC1 [45]. Within the resolution limits of light microscopy, however, we cannot address the question, where Rad21TEV-myc exactly localizes within the SC. Analysis of the SC in Rad21TEV-myc expressing flies via immuno-EM will help to resolve this issue. Our inability to detect uncleaved Rad21TEV-myc localizing to meiotic chromatin in whole mount preparations may be due to epitope masking by a component, which might have been lost during the extensive washing steps with detergent-containing buffer in the chromosome squash preparations.
Taken together, we put forward a model in which at least two types of cohesin complexes are required during Drosophila oogenesis. Firstly, cohesin connecting the chromosome cores to the components of the SC contains the α-kleisin Rad21, possibly composed of multiple interconnected rings. Secondly, cohesin complexes holding together sister chromatids, either contain a very loosely conserved α-kleisin, which awaits to be discovered, or one of the non-kleisin cohesion proteins SOLO or ORD. In the latter case, it will be interesting to find out whether these proteins are substrates of separase.
Materials and Methods
Drosophila stocks and transgene construction
Flies expressing variants of Rad21, which are TEV protease cleavable and C-terminally fused to ten copies of the human c-myc epitope tag, in a Rad21 mutant background, have been described [44]. Expression of these variants is driven by the ubiquitously active α-tubulin 84B promoter. The nanos-GAL4 driver line (y1 w*; P{w[+mC] = GAL4-nos.NGT}40), the maternal triple driver (MTD-GAL4) [55], as well as the c(2)MEP2115 stock [3], [39] were obtained from the Drosophila stock center (Bloomington, Indiana). The maternal alpha-tubulin GAL4 (mat-GAL4) driver line has been described previously [56]. As source for TEV protease, we constructed transgenes encoding a modified enzyme (NLS-V5-TEVS219V), which possesses an N-terminal nuclear localization signal (NLS) followed by a V5 epitope tag and a valine instead of a serine residue at position 219, resulting in inhibition of self-cleavage and in about twofold higher activity levels [57]. We have exclusively used NLS-V5-TEVS219V in this study and, for simplicity, refer to it as TEV protease throughout. To allow TEV protease expression during oogenesis, the NLS-V5-TEVS219V coding sequence was cloned into pUASP1 [58]. Transgenic strains were established after injection into w1 embryos using established procedures.
To obtain flies expressing a C(2)M variant tagged at its C-terminus with six copies of the hemagglutinin tag (6×HA) under control of the c(2)m genomic regulatory sequences, we replaced the 10×myc tag in a progenitor plasmid of the construct pCaSpeR-gC(2)M-myc [39] by the coding sequence for 6×HA. Briefly, a BamHI-XbaI-fragment containing the 3′-terminal part of c(2)m including the 10×myc encoding sequence was subcloned into pBSSK+ (Stratagene). An AgeI site was introduced immediately upstream the c(2)m stop codon by inverse PCR using the oligonucleotides C(2)M7 (5′- GGTGAGACCGGTTGAATATTTTTAGATAATTTTTTTCAAG-3′) and C(2)M8 (5′-CGTTCAACCGGTCTCACTCAGCATAAGATTG-3′) to yield pBSSK+ - BamHI-C(2)M-(AgeI)-XbaI. This step also removed the sequence encoding 10×myc. Next, an XhoI-BamHI fragment containing the 5′-terminal region of c(2)m including flanking genomic sequences was cloned into pBSSK+-BamHI-C(2)M-(AgeI)-XbaI resulting in pBSSK+-gC(2)M-(AgeI). The sequence encoding the 6×HA tag was obtained from the plasmid pUASP-HA-Sse [59] and cloned into the unique AgeI site of pBSSK+-gC(2)M-(AgeI). Finally, the complete insert was transferred as a 4.2 kb NotI-Asp718 fragment into the pattB vector [60]. Transgenic lines were generated by germline transformation of pattB-gC(2)M-HA into y1, w1, M[vas-int]ZH2A; M[3x3P-RFP,attP']ZH51D embryos [60].
To obtain lines carrying a functional Rad21-EGFP transgene, a construct similar to Rad21-10myc was generated. Briefly, the EGFP coding sequence was amplified using the primers SH257 (5′-CGTCTGTTCGAAAACCCAAAAATTGGCGGCGGCATGGTGAGCAAGG-3′) and SH258 (5′-CGTCTGTTCGAACTACTTGTACAGCTCGTCCATGC-3′) and cloned into the naturally occurring BstBI-site upstream of the Rad21 translational stop codon in the Rad21 cDNA clone LD14219 (BDGP). After introducing an additional Acc65I site in the polylinker upstream of the Rad21 coding sequence, the complete Rad21-EGFP fragment was cloned as an Acc65I fragment into the modified pCaSpeR vector used for generating Rad21TEV-myc lines [44]. This vector allows expression of genes inserted in the unique Acc65I site under control of the ubiquitous active α-tubulin 84B promoter. Transgenic lines were established after P-element mediated germ-line transformation using pCaSpeR{w+, αtub-Rad21-EGFP} and injection into embryos derived from parents with the genotype Rad21ex3/TM3, Ser.
For the construction of a putative separase-resistant variant of Rad21, Rad21NC-myc, we employed a PCR-based strategy to exchange the codons 175 and 474 specifying arginines within the separase consensus sequences EXXR into codons specifying alanine residues. We have chosen these two sites because they align well with the known separase cleavage sites in Scc1/Rad21 from humans and yeasts [23]. As template for the PCR reactions, the Rad21 cDNA-based plasmid clone pUAS-Rad21-10myc [44] was used. A first 625 bp fragment comprising the Rad21 5′-UTR up to the region encoding the mutated first separase consensus cleavage site (EIIA) was PCR-amplified using the primers SH341 (ATAAGGCCGGCCACGAGACAGTTTTAGGTGATG) and SH342 (GAAGGTATACTGCAGGCTATAATTTCAGGCGTTTCTGC). A second 910 bp fragment corresponding to the Rad21 region from the mutated first putative separase cleavage site (EIIA) up to the mutated second separase consensus cleavage site (EVLA) was PCR-amplified using the primers SH343 (TTATAGCCTGCAGTATACCTTCAAATATTAATGATAAAA) and SH344 (TTCGCAGCTAGCACTTCCGGAGCTTCCAAACT). A third 1208 bp fragment corresponding to the Rad21 region from the mutated second separase consensus cleavage site (EVLA), through the C-terminal fused c-myc tag, was PCR-amplified using the primers SH345 (GGAAGTGCTAGCTGCGAATCATAAATCTCTAGGG) and SH346 (GTAGGCGCGCCATTAAAACAGATTTACATTCAACTT). The three PCR-generated DNA fragments partially overlap in the regions encoding the mutated separase consensus cleavage sites. After purification using the PCR purification kit (Thermo Scientific), the three PCR products were pooled and served as template for a final PCR using the flanking primers SH341 and SH346. The final 2696 bp PCR-product was cloned as an FseI/AscI fragment into a modified pUASP-vector containing unique FseI and AscI sites within its multiple cloning site. Transgenic lines were generated by P-element mediated germline transformation of w1-embryos using established procedures. For expression of this Rad21 variant in the developing eye, the ey-GAL4 [61] driver line was used and for expression during oogenesis the MTD-GAL4 driver line [55].
For the construction of the transgene P{w+, UASP-NSlmb-vhh-GFP4}, the EcoRI – XbaI insert fragment was isolated from P{w+, UAST-NSlmb-vhh-GFP4} [46], and inserted into the corresponding sites of pUASP1 [58]. Transgenic lines were generated by P-element mediated germline transformation of w1-embryos using established procedures.
For deGradFP dependent destruction of Rad21-EGFP during oogenesis, we generated w*; P{w+, UASP-NSlmb-vhh-GFP4}II.1/nos-GAL4; Rad21ex3, P{w+, αtub-Rad21-EGFP} III.1/Rad21ex3, P{w+, αtub-Rad21-EGFP} III.1 females by standard crossing schemes. As controls, we also generated w*; P{w+, UASP-NSlmb-vhh-GFP4}II.1/nos-GAL4; Rad21ex3, P{w+, αtub-Rad21-EGFP}III.1/+ as well as w*; P{w+, UASP-NSlmb-vhh-GFP4}II.1/+; Rad21ex3, P{w+, αtub-Rad21-EGFP}III.1/Rad21ex3, P{w+, αtub-Rad21-EGFP}III.1 females.
Immunoblotting and immunoprecipitation experiments
For the immunoblotting experiments shown in Fig. 1, ovaries of 4–5 day old females fattened with yeast were dissected in 1× PBS and stage 14 oocytes were isolated and homogenized in SDS gel sample buffer. Protein samples were separated on Tris-glycine based polyacrylamide gels and blotted onto nitrocellulose membranes. For detection of myc epitope tags, HA epitope tags, V5 epitope tags, FLAG epitope tags and α-tubulin, the mouse monoclonal antibodies 9E10 [62], 12CA5 [63], anti-V5 (Invitrogen), anti-FLAG (Sigma) and DM1A (Sigma) were used, respectively. A guinea pig polyclonal antibody against Rad21 [39] and a rabbit antibody against EGFP [64] have been described. For detection of bound antibodies on immunoblots, the horseradish peroxidase based system from p.j.k was used according to the manufacturer's recommendations.
For the immunoprecipitation experiments shown in Fig. 4A, embryos were collected on apple juice agar plates for 1.5 h at 25°C. After dechorionization, the eggs were homogenized in 4× volume of lysis buffer (50 mM HEPES at pH 7.5, 60 mM NaCl, 3 mM MgCl2, 1 mM CaCl2, 0.2% Triton X-100, 0.2% Nonidet NP-40, 10% glycerol, 2 mM Pefabloc, 2 mM Benzamidin, 10 µg/ml Aprotinin, 2 µg/ml Pepstatin A, 10 µg/ml Leupeptin). The extracts were centrifuged and the supernatants were used for immunoprecipitation with anti-HA agarose beads (Roche). After 3 hours incubation at 4°C under rotation, the beads were washed 5× with lysis buffer and transferred into mobicol columns (MoBiTec). Bound proteins were eluted by adding 3× SDS sample buffer (6% SDS, 0.3 M β-mercaptoethanol, 30% glycerol, 0.3% bromophenol blue, 0.15 M Tris/HCl, pH 6.8) and boiling of the sample. The immunoprecipitates as well as samples of the input fractions and supernatants after precipitation were analyzed by immunoblotting.
For the in vitro interaction assays, proteins were synthesized using the TNT SP6 coupled reticulocyte lysate system (Promega) allowing coupled in vitro transcription and translation. To obtain the coding region of the SC components (C(3)G, C(2)M, Corona) and the cohesion proteins (SMC1, ORD, SOLO), RNA from ovaries was isolated and cDNA was synthesized using RevertAid H Minus M-MuLV Reverse Transcriptase (Fermentas) according to the manufacturer's recommendations. As templates for the TNT reactions, the reading frames of the respective genes were cloned into the expression vector pCS2 (F/A) [64] or derivatives thereof, allowing an N-terminal translational fusion with three copies of the FLAG epitope tag or with six copies of the myc epitope tag. For co-expression, equal amounts of plasmid constructs were added to the components of the TNT kit. Synthesized proteins were labeled by incorporation of [35S]methionine. For immunoprecipitation, anti-Flag or anti-myc agarose beads (Sigma) were used.
Immunofluorescence microscopy
Ovaries were dissected in 1× PBS and fixed at room temperature for 20 min in a mixture of 300 µl heptane and 150 µl ovary fixation solution (1× PBS, 0.5% Nonidet NP 40 and 2% para-formaldehyde). Fixed ovaries were blocked for 1 h in PBS containing 0.2% Tween (PBTw) and 10% normal goat serum (NGS). Spread preparations of chromosomes were done as previously described [45]. Rabbit antibodies against Cenp-C [65] and against C(3)G [66] have been described and were used at a 1∶3,000 dilution. For some experiments, we used an anti C(3)G antibody we have raised in guinea pigs by immunization with a bacterially expressed C(3)G fragment corresponding to the C-terminus (aa 565–743). A rat antibody against Cid/Cenp-A (4F8, [67]) was diluted 1∶200. For SMC1 staining, a polyclonal antiserum was raised in rabbits using a bacterially expressed protein fragment corresponding to the N-terminal 133 amino acids of SMC1. The affinity purified antibody was used at a 1∶400 dilution. Antibodies against the HA epitope tag (Roche), the myc epitope tag (Sigma), and the V5 epitope tag (Invitrogen) were used at 1∶10, 1∶10 and 1∶500, respectively. All primary antibodies were diluted in PBTw +10% NGS. After washing twice in PBTw, secondary goat antibodies conjugated with Alexa 488 or Cy3 (Molecular Probes) were applied for 2 h in PBTw containing 5% NGS, followed by additional washes in PBTw. DNA was stained with Hoechst 33258 (1 µg/ml). Fluorescence images were acquired with a Leica SP5 confocal system (Leica Microsystems, Germany) or a Zeiss Axioplan 2 epifluorescence microscope. All images were processed using ImageJ v1.41 (National Institutes of Health, USA). For scoring SC disassembly, we have recorded the stages of those egg chambers showing complete SC disassembly, as indicated by the presence of only dot-like C(3)G signals within the oocyte chromatin and strong C(3)G staining of the oocyte nucleoplasm. The assignment of the stages was done based on size of the egg chambers as determined by equatorial focal planes. Because these planes only rarely allowed illustration of the oocyte nuclei, non-equatorial, and consequently smaller, sections containing the oocyte nuclei are shown in figs. 1, 2, 3, S5 and S7. To assess the significance of the differences in SC disassembly timing (Fig. 3A), we tested pairwise between the respective control situations (no TEV or no NSlmb-vhh-GFP4 expression) and the Rad21 degradation situations (TEV or NSlmb-vhh-GFP4 expression in Rad21TEV-myc, Rad21ex or Rad21-EGFP, Rad21ex background, respectively) using the Mann-Whitney U test (STATISTICA, StatSoft, Tulsa, OK, USA).
Fluorescent in situ hybridization (FISH)
The X chromosome-specific 359 bp repeat was amplified by PCR with Drosophila genomic DNA as template [68]. The PCR product was digested overnight with a mixture of the restriction enzymes AluI, HaeIII, Tru1I, MspI, RsaI, and Sau3AI. Digested DNA was precipitated, dissolved in water, denatured at 100°C for 1 min and chilled on ice. The AATAT repeat specific for chromosome 4 was synthesized as a single-stranded oligonucleotide ((AATAT)6; Metabion international AG, Germany). 3′-Tailing of the single stranded DNAs with the reactive nucleotide Aminoallyl dUTP analog was done by using Terminal deoxynucleotidyl Transferase (Roche) at 37°C for 2 h in a reaction mixture containing 200 mM Na-Cacodylate (pH 7.2), 100 µM DTT, 1 mM CoCl2, 50 µM Aminoallyl dUTP (ARES DNA Alexa Fluor 555/647 labeling kit, Molecular probes) and 5 µM unlabeled dTTP. Reactions were stopped by adding 5 mM EDTA. Aminoallyl-conjugated probes were precipitated, dissolved in water and labeled with Alexa Fluor 555 or Alexa Fluor 647 in labeling buffer for 2 h in the dark, followed by quenching of the reactions with 150 µM hydroxylamine. Labeled probes were precipitated and dissolved in elution buffer.
FISH was done on stage 14 oocytes as described in [69] with some modifications. Oocytes were fixed in heptane/oocyte fixation solution, rinsed three times in 2× SSCT (0.3 M NaCl, 30 mM sodium citrate, 0.1% Tween 20), sequentially washed with 2× SSCT-20% formamide, 2× SSCT-40% formamide, and 2× SSCT-50% formamide for 10 min each followed by incubation in fresh 2× SSCT-50% formamide for 1–2 hrs at 37°C. The oocytes were transferred to 36 µl of hybridization buffer (20% dextrane sulfate, 15% formamide in 2× SSCT) and 100 ng of each fluorescently labelled probe was added. Probe and chromosomal DNA were denatured at 91°C for 2 min and the hybridization reaction was carried out overnight at 37°C. After hybridization, pre-warmed (37°C) 2× SSCT-50% formamide was added to the sample. Oocytes were washed three times with pre-warmed 2× SSCT-50% formamide, once with 2× SSCT-40% formamide, and 2× SSCT-20% formamide for 10 min/wash. Then, the oocytes were washed three times with 2× SSCT for 10 min each, rinsed three times with PBST and treated with Hoechst 33258 (1 µg/ml in PBS) to stain DNA. Finally, the oocytes were washed once with PBS for 5 min and mounted in 70% glycerol, 50 mM Tris-Cl (pH 9.5), 10 mg/ml propyl gallate, 0.5 mg/ml p-phenylenediamine in 1× PBS.
To enrich for oocytes progressing through meiosis II, approximately 300 females fattened for three days on yeast were put in collection cages and after a pre-collection for 1 h at 25°C, eggs were collected every 20 to 40 minutes for 5 hours. The eggs were immediately dechorionized and fixed with Methanol. Eggs from all collections were pooled and subjected to FISH as described above.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. PetronczkiM, SiomosMF, NasmythK (2003) Un menage a quatre: the molecular biology of chromosome segregation in meiosis. Cell 112: 423–440.
2. McKimKS, Green-MarroquinBL, SekelskyJJ, ChinG, SteinbergC, et al. (1998) Meiotic synapsis in the absence of recombination. Science 279: 876–878.
3. ManheimEA, McKimKS (2003) The Synaptonemal complex component C(2)M regulates meiotic crossing over in Drosophila. Curr Biol 13: 276–285.
4. PageSL, HawleyRS (2001) c(3)G encodes a Drosophila synaptonemal complex protein. Genes Dev 15: 3130–3143.
5. OliveiraRA, NasmythK (2010) Getting through anaphase: splitting the sisters and beyond. Biochem Soc Trans 38: 1639–1644.
6. HaeringCH, JessbergerR (2012) Cohesin in determining chromosome architecture. Exp Cell Res 318: 1386–1393.
7. GuacciV, KoshlandD, StrunnikovA (1997) A direct link between sister chromatid cohesion and chromosome condensation revealed through the analysis of MCD1 in S-cerevisiae. Cell 91: 47–57.
8. MichaelisC, CioskR, NasmythK (1997) Cohesins: Chromosomal proteins that prevent premature separation of sister chromatids. Cell 91: 35–45.
9. HaeringCH, FarcasAM, ArumugamP, MetsonJ, NasmythK (2008) The cohesin ring concatenates sister DNA molecules. Nature 454: 297–301.
10. McNicollF, StevenseM, JessbergerR (2013) Cohesin in gametogenesis. Curr Top Dev Biol 102: 1–34.
11. KleinF, MahrP, GalovaM, BuonomoSB, MichaelisC, et al. (1999) A central role for cohesins in sister chromatid cohesion, formation of axial elements, and recombination during yeast meiosis. Cell 98: 91–103.
12. WatanabeY, NurseP (1999) Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature 400: 461–464.
13. Gutierrez-CaballeroC, HerranY, Sanchez-MartinM, SujaJA, BarberoJL, et al. (2011) Identification and molecular characterization of the mammalian alpha-kleisin RAD21L. Cell Cycle 10: 1477–1487.
14. IshiguroK, KimJ, Fujiyama-NakamuraS, KatoS, WatanabeY (2011) A new meiosis-specific cohesin complex implicated in the cohesin code for homologous pairing. EMBO Rep 12: 267–275.
15. LeeJ, HiranoT (2011) RAD21L, a novel cohesin subunit implicated in linking homologous chromosomes in mammalian meiosis. J Cell Biol 192: 263–276.
16. LlanoE, HerranY, Garcia-TunonI, Gutierrez-CaballeroC, de AlavaE, et al. (2012) Meiotic cohesin complexes are essential for the formation of the axial element in mice. J Cell Biol 197: 877–885.
17. HerranY, Gutierrez-CaballeroC, Sanchez-MartinM, HernandezT, VieraA, et al. (2011) The cohesin subunit RAD21L functions in meiotic synapsis and exhibits sexual dimorphism in fertility. EMBO J 30: 3091–3105.
18. BannisterLA, ReinholdtLG, MunroeRJ, SchimentiJC (2004) Positional cloning and characterization of mouse mei8, a disrupted allelle of the meiotic cohesin Rec8. Genesis 40: 184–194.
19. RevenkovaE, EijpeM, HeytingC, HodgesCA, HuntPA, et al. (2004) Cohesin SMC1 beta is required for meiotic chromosome dynamics, sister chromatid cohesion and DNA recombination. Nat Cell Biol 6: 555–562.
20. XuH, BeasleyMD, WarrenWD, van der HorstGT, McKayMJ (2005) Absence of mouse REC8 cohesin promotes synapsis of sister chromatids in meiosis. Dev Cell 8: 949–961.
21. SeversonAF, LingL, van ZuylenV, MeyerBJ (2009) The axial element protein HTP-3 promotes cohesin loading and meiotic axis assembly in C. elegans to implement the meiotic program of chromosome segregation. Genes Dev 23: 1763–1778.
22. UhlmannF, WernicD, PoupartMA, KooninEV, NasmythK (2000) Cleavage of cohesin by the CD clan protease separin triggers anaphase in yeast. Cell 103: 375–386.
23. HaufS, WaizeneggerIC, PetersJM (2001) Cohesin cleavage by separase required for anaphase and cytokinesis in human cells. Science 293: 1320–1323.
24. KudoNR, AngerM, PetersAH, StemmannO, TheusslHC, et al. (2009) Role of cleavage by separase of the Rec8 kleisin subunit of cohesin during mammalian meiosis I. J Cell Sci 122: 2686–2698.
25. KudoNR, WassmannK, AngerM, SchuhM, WirthKG, et al. (2006) Resolution of chiasmata in oocytes requires separase-mediated proteolysis. Cell 126: 135–146.
26. BuonomoSB, ClyneRK, FuchsJ, LoidlJ, UhlmannF, et al. (2000) Disjunction of homologous chromosomes in meiosis I depends on proteolytic cleavage of the meiotic cohesin Rec8 by separin. Cell 103: 387–398.
27. KitajimaTS, MiyazakiY, YamamotoM, WatanabeY (2003) Rec8 cleavage by separase is required for meiotic nuclear divisions in fission yeast. EMBO J 22: 5643–5653.
28. KitajimaTS, SakunoT, IshiguroK, IemuraS, NatsumeT, et al. (2006) Shugoshin collaborates with protein phosphatase 2A to protect cohesin. Nature 441: 46–52.
29. RiedelCG, KatisVL, KatouY, MoriS, ItohT, et al. (2006) Protein phosphatase 2A protects centromeric sister chromatid cohesion during meiosis I. Nature 441: 53–61.
30. LeeJ, YokotaT, YamashitaM (2002) Analyses of mRNA expression patterns of cohesin subunits Rad21 and Rec8 in mice: germ cell-specific expression of rec8 mRNA in both male and female mice. Zoolog Sci 19: 539–544.
31. ParraMT, VieraA, GomezR, PageJ, BenaventeR, et al. (2004) Involvement of the cohesin Rad21 and SCP3 in monopolar attachment of sister kinetochores during mouse meiosis I. J Cell Sci 117: 1221–1234.
32. XuH, BeasleyM, VerschoorS, InselmanA, HandelMA, et al. (2004) A new role for the mitotic RAD21/SCC1 cohesin in meiotic chromosome cohesion and segregation in the mouse. EMBO Rep 5: 378–384.
33. PrietoI, TeaseC, PezziN, BuesaJM, OrtegaS, et al. (2004) Cohesin component dynamics during meiotic prophase I in mammalian oocytes. Chromosome Res 12: 197–213.
34. GomezR, ValdeolmillosA, ParraMT, VieraA, CarreiroC, et al. (2007) Mammalian SGO2 appears at the inner centromere domain and redistributes depending on tension across centromeres during meiosis II and mitosis. EMBO Rep 8: 173–180.
35. EijpeM, OffenbergH, JessbergerR, RevenkovaE, HeytingC (2003) Meiotic cohesin REC8 marks the axial elements of rat synaptonemal complexes before cohesins SMC1beta and SMC3. J Cell Biol 160: 657–670.
36. PrietoI, PezziN, BuesaJM, KremerL, BarthelemyI, et al. (2002) STAG2 and Rad21 mammalian mitotic cohesins are implicated in meiosis. EMBO Rep 3: 543–550.
37. Tachibana-KonwalskiK, GodwinJ, van der WeydenL, ChampionL, KudoNR, et al. (2010) Rec8-containing cohesin maintains bivalents without turnover during the growing phase of mouse oocytes. Genes Dev 24: 2505–2516.
38. SchleifferA, KaitnaS, Maurer-StrohS, GlotzerM, NasmythK, et al. (2003) Kleisins: a superfamily of bacterial and eukaryotic SMC protein partners. Mol Cell 11: 571–575.
39. HeidmannD, HornS, HeidmannS, SchleifferA, NasmythK, et al. (2004) The Drosophila meiotic kleisin C(2)M functions before the meiotic divisions. Chromosoma 113: 177–187.
40. BickelSE, WymanDW, Orr-WeaverTL (1997) Mutational analysis of the Drosophila sister-chromatid cohesion protein ORD and its role in the maintenance of centromeric cohesion. Genetics 146: 1319–1331.
41. YanR, McKeeBD (2013) The cohesion protein SOLO associates with SMC1 and is required for synapsis, recombination, homolog bias and cohesion and pairing of centromeres in Drosophila Meiosis. PLoS Genet 9: e1003637.
42. YanR, ThomasSE, TsaiJH, YamadaY, McKeeBD (2010) SOLO: a meiotic protein required for centromere cohesion, coorientation, and SMC1 localization in Drosophila melanogaster. J Cell Biol 188: 335–349.
43. WebberHA, HowardL, BickelSE (2004) The cohesion protein ORD is required for homologue bias during meiotic recombination. J Cell Biol 164: 819–829.
44. PauliA, AlthoffF, OliveiraRA, HeidmannS, SchuldinerO, et al. (2008) Cell-type-specific TEV protease cleavage reveals cohesin functions in Drosophila neurons. Dev Cell 14: 239–251.
45. KhetaniRS, BickelSE (2007) Regulation of meiotic cohesion and chromosome core morphogenesis during pachytene in Drosophila oocytes. J Cell Sci 120: 3123–3137.
46. CaussinusE, KancaO, AffolterM (2012) Fluorescent fusion protein knockout mediated by anti-GFP nanobody. Nat Struct Mol Biol 19: 117–121.
47. TakeoS, LakeCM, Morais-de-SaE, SunkelCE, HawleyRS (2011) Synaptonemal complex-dependent centromeric clustering and the initiation of synapsis in Drosophila oocytes. Curr Biol 21: 1845–1851.
48. BickelSE, Orr-WeaverTL, BalickyEM (2002) The sister-chromatid cohesion protein ORD is required for chiasma maintenance in Drosophila oocytes. Curr Biol 12: 925–929.
49. Howard-TillRA, LukaszewiczA, NovatchkovaM, LoidlJ (2013) A single cohesin complex performs mitotic and meiotic functions in the protist tetrahymena. PLoS Genet 9: e1003418.
50. Tachibana-KonwalskiK, GodwinJ, BorsosM, RattaniA, AdamsDJ, et al. (2013) Spindle assembly checkpoint of oocytes depends on a kinetochore structure determined by cohesin in meiosis I. Curr Biol 23: 2534–2539.
51. MurdochB, OwenN, StevenseM, SmithH, NagaokaS, et al. (2013) Altered cohesin gene dosage affects Mammalian meiotic chromosome structure and behavior. PLoS Genet 9: e1003241.
52. MolnarM, BahlerJ, SipiczkiM, KohliJ (1995) The rec8 gene of Schizosaccharomyces pombe is involved in linear element formation, chromosome pairing and sister-chromatid cohesion during meiosis. Genetics 141: 61–73.
53. ZhangN, KuznetsovSG, SharanSK, LiK, RaoPH, et al. (2008) A handcuff model for the cohesin complex. J Cell Biol 183: 1019–1031.
54. AndersonLK, RoyerSM, PageSL, McKimKS, LaiA, et al. (2005) Juxtaposition of C(2)M and the transverse filament protein C(3)G within the central region of Drosophila synaptonemal complex. Proc Natl Acad Sci U S A 102: 4482–4487.
55. PetrellaLN, Smith-LeikerT, CooleyL (2007) The Ovhts polyprotein is cleaved to produce fusome and ring canal proteins required for Drosophila oogenesis. Development 134: 703–712.
56. MicklemDR, DasguptaR, ElliottH, GergelyF, DavidsonC, et al. (1997) The mago nashi gene is required for the polarisation of the oocyte and the formation of perpendicular axes in Drosophila. Curr Biol 7: 468–478.
57. KapustRB, TozserJ, FoxJD, AndersonDE, CherryS, et al. (2001) Tobacco etch virus protease: mechanism of autolysis and rational design of stable mutants with wild-type catalytic proficiency. Protein Eng 14: 993–1000.
58. JägerH, RauchM, HeidmannS (2005) The Drosophila melanogaster condensin subunit Cap-G interacts with the centromere-specific histone H3 variant CID. Chromosoma 113: 350–361.
59. JägerH, HerzigA, LehnerCF, HeidmannS (2001) Drosophila Separase is required for sister chromatid separation and binds to PIM and THR. Genes Dev 15: 2572–2584.
60. BischofJ, MaedaRK, HedigerM, KarchF, BaslerK (2007) An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc Natl Acad Sci U S A 104: 3312–3317.
61. HazelettDJ, BourouisM, WalldorfU, TreismanJE (1998) decapentaplegic and wingless are regulated by eyes absent and eyegone and interact to direct the pattern of retinal differentiation in the eye disc. Development 125: 3741–3751.
62. EvanGI, LewisGK, RamsayG, BishopJM (1985) Isolation of monoclonal antibodies specific for human c-myc proto-oncogene product. Mol Cell Biol 5: 3610–3616.
63. NimanHL, HoughtenRA, WalkerLE, ReisfeldRA, WilsonIA, et al. (1983) Generation of protein-reactive antibodies by short peptides is an event of high frequency: implications for the structural basis of immune recognition. Proc Natl Acad Sci USA 80: 4949–4953.
64. HerzogS, Nagarkar JaiswalS, UrbanE, RiemerA, FischerS, et al. (2013) Functional dissection of the Drosophila melanogaster condensin subunit Cap-G reveals its exclusive association with condensin I. PLoS Genet 9: e1003463.
65. HeegerS, LeismannO, SchittenhelmR, SchraidtO, HeidmannS, et al. (2005) Genetic interactions of separase regulatory subunits reveal the diverged Drosophila Cenp-C homolog. Genes Dev 19: 2041–2053.
66. HongA, Lee-KongS, IidaT, SugimuraI, LillyMA (2003) The p27cip/kip ortholog dacapo maintains the Drosophila oocyte in prophase of meiosis I. Development 130: 1235–1242.
67. PadekenJ, MendiburoMJ, ChlamydasS, SchwarzHJ, KremmerE, et al. (2013) The nucleoplasmin homolog NLP mediates centromere clustering and anchoring to the nucleolus. Mol Cell 50: 236–249.
68. HsiehT, BrutlagD (1979) Sequence and sequence variation within the 1.688 g/cm3 satellite DNA of Drosophila melanogaster. J Mol Biol 135: 465–481.
69. DernburgAF, BromanKW, FungJC, MarshallWF, PhilipsJ, et al. (1996) Perturbation of nuclear architecture by long-distance chromosome interactions. Cell 85: 745–759.
Štítky
Genetika Reprodukčná medicínaČlánok vyšiel v časopise
PLOS Genetics
2014 Číslo 8
- Je „freeze-all“ pro všechny? Odborníci na fertilitu diskutovali na virtuálním summitu
- Gynekologové a odborníci na reprodukční medicínu se sejdou na prvním virtuálním summitu
Najčítanejšie v tomto čísle
- Meta-Analysis of Genome-Wide Association Studies in African Americans Provides Insights into the Genetic Architecture of Type 2 Diabetes
- KDM6 Demethylase Independent Loss of Histone H3 Lysine 27 Trimethylation during Early Embryonic Development
- The RNA Helicases AtMTR4 and HEN2 Target Specific Subsets of Nuclear Transcripts for Degradation by the Nuclear Exosome in
- EF-P Dependent Pauses Integrate Proximal and Distal Signals during Translation