
The Impact of Population Demography and Selection on the Genetic Architecture of Complex Traits
Many human populations have dramatically expanded over the last several thousand years. I use population genetic models to investigate how recent population expansions affect patterns of mutations that reduce reproductive fitness and contribute to the genetic basis of complex traits (including common disease). I show that recent population growth increases the proportion of mutations found in the population that reduce fitness. When mutations that have the greatest effect on reproductive fitness also have the greatest effect on a complex trait, more of the heritability of the trait is due to mutations at very low-frequency in populations that have recently expanded, as compared to populations that have not. Also, under this model, for a given sample size and false-positive rate, fewer variants show statistically significant associations with the trait in the population that has expanded than in one that has not. Both of these findings suggest that recent population growth may make it more difficult to fully elucidate the genetic basis of complex traits that are directly or indirectly correlated with reproductive fitness.
Published in the journal:
The Impact of Population Demography and Selection on the Genetic Architecture of Complex Traits. PLoS Genet 10(5): e32767. doi:10.1371/journal.pgen.1004379
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1004379
Summary
Many human populations have dramatically expanded over the last several thousand years. I use population genetic models to investigate how recent population expansions affect patterns of mutations that reduce reproductive fitness and contribute to the genetic basis of complex traits (including common disease). I show that recent population growth increases the proportion of mutations found in the population that reduce fitness. When mutations that have the greatest effect on reproductive fitness also have the greatest effect on a complex trait, more of the heritability of the trait is due to mutations at very low-frequency in populations that have recently expanded, as compared to populations that have not. Also, under this model, for a given sample size and false-positive rate, fewer variants show statistically significant associations with the trait in the population that has expanded than in one that has not. Both of these findings suggest that recent population growth may make it more difficult to fully elucidate the genetic basis of complex traits that are directly or indirectly correlated with reproductive fitness.
Introduction
Genome-wide association studies (GWAS) have successfully detected associations between hundreds of common single nucleotide polymorphisms (SNPs) and complex traits in humans [1], [2]. While this catalog of genes has revealed important biological insights, for most traits the discovered associations can account only for a small fraction of the heritability measured from family-based studies [3]. This difference in the heritability observed in familial studies and the heritability explained by associated SNPs has been termed “missing heritability,” and there is tremendous interest in the human genetics community to find it [3]–[5].
One possibility that has received particular attention is that the missing heritability lies in rare variants that have large effect sizes [3]. Because a risk variant is rare in the population, an association between the variant and the phenotypes of interest may not have been detected using traditional GWAS. Instead, at present, such variants must be assayed through direct sequencing. Due to technological advances (e.g. next-generation sequencing) [6], [7], combined with newer analytical methods designed for analyzing full sequence data [8]–[11], exome and full genome-sequencing studies are now being implemented in human genetics. The progression to sequencing data has already proved fruitful for the identification of causal mutations for several Mendelian diseases [6], [12]–[16]. Full sequence data [17], [18] is starting to reveal a richer picture of low-frequency genetic variation (minor allele frequency <0.5%), which may, in turn, increase the community's ability to implicate rare variants in risk of complex disease [4], [19]–[25]. Further, such studies should allow researchers to empirically determine the extent to which rare variants account for the missing heritability of complex traits [26]–[30]. However, before these new technological and methodological advances can reach their full potential, a more thorough understanding of low-frequency genetic variation in multiple human populations is essential.
To learn about patterns of rare genetic variation, several studies have sequenced hundreds of genes or complete exomes in thousands of individuals [31]–[34]. These studies have made two important discoveries. First, they have found a larger number of rare variants than was expected under previous models of human population history. It has been argued that this excess of rare variants can be explained by the recent explosion in human population size [31], [32], [35]–[37]. Second, these studies have documented a plethora of rare, nonsynonymous SNPs that are likely evolutionarily deleterious and may be of medical relevance.
Comparatively less work has been done, however, to examine the implications that recent population history has had on the architecture of complex traits (but see the recent paper by Simons et al. [38]). It is unclear whether population history, and recent population growth in particular, affects the number, frequency, and effect sizes of mutations that contribute risk to complex traits. Addressing this question is critical for finding the “missing heritability” in different populations, as well as performing the most powerful association studies to implicate specific variants in disease risk. Over a decade ago, it was recognized that the power to associate common variants with complex disease varied across populations [39]–[41]. This was largely due to asymmetry in the extent of linkage disequilibrium (LD) across populations as a result of differences in demographic history [42]–[44]. While the issue of LD is less relevant when considering rare variants, the topic of population choice for association studies has received substantially less attention when considering rare variants, despite its potential importance.
Here I use population genetic models to investigate the effect of recent population growth on patterns of deleterious genetic variation, the architecture of complex traits, and the ability to associate causal variants with the trait in models where a proportion of the trait's heritability is accounted for by mutations in a subset of the exome. Specifically, I show that recent population growth increases the input of deleterious mutations into the population, directly causing a proportional excess of deleterious genetic variation segregating in the population. Second, if a mutation's effect on reproductive fitness is correlated with its effect on a complex trait (such as a disease), I show that recent population growth increases the amount of the additive genetic variance of the trait that is accounted for by low-frequency variants relative to that in a population that did not expand recently. Further, I demonstrate that recent population growth leads to an increase in the number of alleles that contribute to the trait relative to what is expected in a population that did not recently expand. Finally, recent population growth decreases the number of SNPs that are significantly associated with the trait, relative to the number detected in a population that did not recently expand. This work indicates that in certain circumstances, recent population history will play an important role in determining the genetic architecture of complex traits in a particular population under study. As such, recent population history is a factor that should be considered when designing and interpreting re-sequencing studies for complex traits.
Methods
Models of population history
I explore several of models of population history (Fig. 1). Because many studies have inferred a population bottleneck in non-African human populations associated with the Out-of-Africa migration process [45]–[50], the first model includes a brief, but severe, reduction in population size (Fig. 1A). After the bottleneck, the population returns to the same size as the ancestral population. This model is referred to as “BN” throughout the rest of paper. The second model of population history also includes the same Out-of-Africa population bottleneck, but now includes an instantaneous, 100-fold population expansion in the last 80 generations, or the last 2000 years, assuming 25 years/generation (Fig. 1B) [51]. This recent explosion in effective population size is meant to approximate the expansion detected in the archeological and historical records as well as in studies of genetic variation [31], [35], [36], [52]. This model is referred to as “BN+growth” throughout the paper. Finally, for comparison purposes, I also investigate a model where a population experienced an ancient 2-fold expansion (Fig. 1C). Such a model is meant to reflect the history of African populations [46], [47], [53] and is referred to as “Old growth” in the paper.

Forward simulations
All results were obtained using the forward-in-time population genetic program described in Lohmueller et al. [54], with minor modifications. Briefly, the program assumes a Wright-Fisher model of population history. Each generation, alleles change frequency stochastically based on binomial sampling and deterministically based on the standard selection equations. Also in each generation, a Poisson distributed number of new mutations enter the population at rate , where . Here is the population size in the ith epoch of population history (see below) and is the per-chromosome per generation mutation rate across all the coding sequence that was simulated. I set for synonymous sites. For nonsynonymous sites, is 2.5 times higher, because of the larger number of sites that, when mutated, give rise to a nonsynonymous mutation. Selection coefficients for new mutations are drawn from a gamma distribution with the parameters as inferred in Boyko et al. [46]. As done in the Poisson Random Field framework, all mutations are assumed to be independent of each other [55].
Step-wise population size changes are included in the model by changing the population size (N) at particular time points. Size changes affect the number of mutations that enter the population during each individual epoch and the magnitude of genetic drift.
For computational efficiency, I divided the population size by 2 and rescaled all times to be two-fold smaller than under the specified model. However, I keep the population scaled mutation rate (), and the population scaled selection coefficient (), equal to the same values as for the larger population. This rescaling is possible because, in the diffusion limit, patterns of genetic variation only depend on the scaled parameters. Such a rescaling is customary in other forward simulation programs [56], [57]. Samples of 1000 chromosomes are taken from the population at different time points to calculate how diversity statistics change over time.
Models of complex traits
To evaluate the effect of recent demographic history on the architecture of complex traits, I simulate individuals who have a quantitative trait. I assume that deleterious (nonsynonymous) mutations in a given mutational target account for some of the heritability of the trait. Such a model is implicitly assumed in exome re-sequencing studies used to implicate rare variants in disease risk. This quantitative trait could represent a trait that is measured on a quantitative scale (e.g. lipid levels) or represent the underlying risk to a dichotomous phenotype (e.g. diabetes). Below I provide a description of the model and parameters.
I investigate models where mutations at a subset of nonsynonymous sites can account for 5%, 10% or, 30% of the variance of the phenotype (i.e. the heritability accounted for by these variants is 5%, 10% or 30%). Thus, the models considered here assume that some fraction of the total heritability of the trait is accounted for by variants within the mutational target (i.e. a portion of the exome) while the rest is accounted for by variants not modeled here (i.e. noncoding portions of the genome). The mutational target size, M, is the number of nonsynonymous sites in the genome that, if mutated, would generate a variant that affects the phenotype. Assuming a mutation rate of 1×10−8 per site per generation, I investigate mutational target sizes of 70 kb and 140 kb. To gain a sense of how these sites could be partitioned into genes, the median length of coding regions of human genes is 1335 bp [58]. Thus, a random gene would have approximately 934 nonsynonymous sites (assuming 70% of the coding sites are nonsynonymous). If all nonsynonymous sites within the gene would, if mutated, produce a causal variant, then the mutational target size of 70 kb would correspond to 75 distinct genes accounting for the specified heritability, and the target size of 140 kb would correspond to 150 distinct causal genes accounting for the heritability. If only half the nonsynonymous sites could be mutated to causal variants, then the number of genes would increase by a factor of 2. In practice, this model is implemented by taking a subset of the nonsynonymous SNPs simulated as described above and then assigning them effects on the trait.
To assign an effect on the trait to a given causal SNP, I follow the model described by Eyre-Walker [59], with modifications described below. Essentially, the ith SNP's effect on a trait, αi, is given by


I then assign trait values (Yj) to each simulated individual. This is done using an additive model,

Results
Recent growth and deleterious variation
First I assess the effect that different population histories (BN, BN+growth, and Old Growth) have on neutral and deleterious genetic variation. Fig. 2A and Fig. 2B show how the number of synonymous and nonsynonymous SNPs, respectively, segregating in a sample of 1000 chromosomes change over time as the simulated populations change in size. The population bottleneck 2000 generations ago resulted in a decrease in the number of SNPs segregating in the BN and BN+growth populations (orange and green lines in Fig. 2A and Fig. 2B). When the populations recovered from the bottleneck and increased in size, the number of SNPs in the population also increased. This increase in the number of SNPs after the recovery from the bottleneck is due to two factors. First, the larger population size allows more new mutations to enter the population. Second, genetic drift has a weaker effect when the population size is large. As such, more SNPs are maintained in the population. The recent explosion in population size (dashed green lines in Fig. 2A and Fig. 2B; BN+growth) rapidly results in a substantial increase in the number of both synonymous and nonsynonymous SNPs segregating in the population. This is due to the extreme increase in the population mutation rate (typically referred to as ) due to the larger population size. Ancient population growth also resulted in an increase in the number of synonymous and nonsynonymous SNPs segregating in the population, via the same mechanisms (purple line in Fig. 2A and Fig 2B; Old growth).

Fig. 2C shows how the proportion of nonsynonymous SNPs segregating in the population changes over time. When populations BN and BN+growth decreased in size during the bottleneck, the proportion of nonsynonymous SNPs in the population also dropped (orange and green lines in Fig. 2C). The reason for this is that, when the population size decreases, rare variants are preferentially lost over common variants. More nonsynonymous than synonymous SNPs are rare, and, as such, the crash in population size results in the loss of more nonsynonymous SNPs than synonymous SNPs. After the population recovers from the bottleneck, the proportion of nonsynonymous SNPs found in the population increases (Fig. 2C). The reason for this increase is that, due to the increase in population size, many new mutations enter the population after the recovery from the bottleneck. Most of these new mutations are nonsynonymous, due to there being more possible nonsynonymous changes than synonymous changes in coding regions. In fact, the proportion of nonsynonymous SNPs segregating in the population immediately after the recovery of the bottleneck is actually higher than that in the ancestral population (Fig. S2). The very recent increase in size in the BN+growth population also results in an increase in the proportion of nonsynonymous SNPs (green line in Fig. 2C). 54.8% of the SNPs in BN+growth are nonsynonymous (green line in Fig. 2C), compared to 52.8% in the BN population (orange line in Fig. 2C). It will take approximately 4Ne (where Ne is the current effective population size) generations for the proportion of deleterious SNPs to reach the equilibrium value for the larger population size (Text S1). The population that underwent an ancient expansion (dotted purple line in Fig. 2C) also experienced an initial increase in the proportion of nonsynonymous SNPs segregating immediately after the expansion. However, because the expansion occurred long ago, selection has had sufficient time to remove many of the nonsynonymous SNPs and bring the proportion of nonsynonymous SNPs in the population below the value seen in the bottlenecked populations, consistent with previous simulations and empirical observations [54]. These results suggest that recent, extreme changes in demographic history can have an impact on patterns of deleterious mutations that are segregating in the population. This pattern also holds with other magnitudes of population growth (Text S1).
Next I examine the average fitness effect of nonsynonymous SNPs segregating in a sample of 1000 chromosomes at different time points in the simulations (Fig. 2D). During the bottleneck, the average segregating SNP in the BN and BN+growth populations is less deleterious than in the ancestral population (orange and green lines in Fig. 2D). This is due to many rare, deleterious SNPs being eliminated from the population as well as fewer new deleterious SNPs entering the population when it is small in size. After the population recovers from the bottleneck, the average segregating SNP became more deleterious. In the first few generations after the recovery, the average SNP is even more deleterious than in the ancestral population. This is due to the increase in the input of deleterious mutations immediately after the recovery from the bottleneck. After a few generations however, negative natural selection has eliminated many, though certainly not all, of these deleterious SNPs from the population. In fact, Fig. 2D shows that even in the present day, the average SNP is more deleterious than that in the ancestral population. This same effect applies even more strongly to the recent population growth within the last 80 generations. Immediately, after growth, the average SNP segregating in the BN+growth population was more strongly deleterious (Fig. 2D) than what is expected in a population that has not expanded. However, during the last 40 generations, selection has eliminated many of the most deleterious SNPs from the population. In the present day, the average SNP in the BN+growth population (green line) is slightly less deleterious than in the BN population (orange line, also see Text S1), consistent with the results of Gazave et al. [65]. This effect is less pronounced with decreasing amounts of population growth (Text S1).
The description of the average strength of selection on a SNP described above does not take into account the frequency of the deleterious SNP in the population. The genetic load, however, does by weighting the selection coefficient by the SNP's frequency [66]. Genetic load is the reduction in mean fitness of the population due to deleterious mutations [67]. I find that, unlike the average selection coefficient, the genetic load is not affected by the demographic history of the population (Fig. S3). Similar results have recently been reported by Simons et al. [38]. Thus, while the recent population growth increases the number of deleterious SNPs segregating in the population, this increase in load is offset by the fact that most of these new deleterious mutations are kept at very low frequency in the population. Put another way, while the load appears to be the same across demographic models, the way in which the populations arrive at that load differs across demographies. The BN+growth population contains many rare deleterious mutations. The BN population contains fewer deleterious mutations, but those that are there tend to be at higher frequencies.
Models of a complex trait that are compatible with empirical results
One important question is to what extent recent population growth affects the architecture of complex traits and our ability to map the genes responsible for disease risk. To investigate this issue, I simulate quantitative phenotypes for individuals sampled from the simulations under the three different demographic models. I investigate two different models for the relationship between a mutation's effect on fitness (the selection coefficient), and its effect on the trait [59]. First, I assume that a mutation's effect on fitness is related to its effect on the trait (τ = 0.5). Here, those mutations that are strongly deleterious have a greater effect on the trait. Second, I investigate a model where a SNP's effect on fitness is independent of its effect on the trait (τ = 0). I also investigate different mutational target sizes (M = 70 kb and M = 140 kb) and the amount of the heritability accounted for by variants occurring within the mutational target (; see Methods; see Discussion for further justification of these models).
Because the model parameters were chosen to reflect what might be observed in exome resequencing data, I test the validity of these parameter values by comparing the expected number of single SNPs associated with a trait to what has been observed in exome sequencing studies. A recent exome sequencing study for type 2 diabetes with 1000 cases and 1000 controls (the same sample size simulated here) reported zero single SNP associations at a 1×10−5 significance level [30]. This result is broadly consistent with models where τ = 0.5 and or and models with τ = 0 and (Fig. S4). Next, an exome sequencing study for schizophrenia with 2,536 cases and 2,543 controls [68] found only common variants with P<10−5, which, while not directly comparable to the sample size used in the present study, is again consistent with models where τ = 0.5 and or and models with τ = 0 and (Fig. S4). Finally, an exome sequencing study for lipids included 2,005 individuals and found two variants with P<10−5 in a single-marker analysis [69]. These results are compatible with a variety of models simulated here (Fig. S4), though Lange et al.'s quantitative association test should have more power than the dichotomous one used here, making precise comparisons more difficult.
Due to the limited number of exome sequencing studies available, I also assess whether my models generate simulated datasets that are compatible with observations from GWAS studies. For simplicity, I make the assumption that the causal variants themselves have been directly assayed or have been imputed through LD with tag SNPs included in the GWAS. This of course is unlikely to be true in practice, particularly for rare variants [17], [18], [70]. Nevertheless, this comparison still serves as a useful benchmark to exclude models that are obviously not consistent with the observed GWAS data, with the caveat that some models that appear inconsistent with the GWAS data may actually fit better if LD and ascertainment bias are properly taken into account.
First, it has been consistently shown that the top SNPs identified through GWAS account for only a very limited amount of the phenotypic variance (often <10%) [3], [71]. I assess the amount of phenotypic variance (VP) explained by the top 50 SNPs that account for the most variance (Table S2) within each simulation replicate. Models where and M = 70 kb or 140 kb predict that the top 50 SNPs account for roughly 30% of the VP. This amount appears to be too high to be compatible with most of the GWAS results if one accepts the premise that GWAS would have detected the variants that explain such a large proportion of the phenotypic variance. However, a model with predicts that the top 50 SNPs will account for about 5% of the phenotypic variance. Such a model cannot be rejected from the currently available GWAS data.
Next, GWAS suggest that most risk loci for complex traits have very small effect sizes and are difficult to detect in samples of only 1000 cases and controls [71]. Table S3 shows the expected number of GWAS hits (P<5×10−8) expected in each simulation replicate in samples of 1000 cases and 1000 controls for the different models of , M, and τ. Models where and M = 70 kb or 140 kb predict that 1–4 significant GWAS hits should be observed. This is too many to be compatible with the observed data. Models with the mutational target accounting for less of the heritability ( and ) predict <1 significant association. Thus, these models cannot be rejected based on GWAS data.
In summary, it is unclear how much of the heritability of complex traits in humans is accounted for by the exome and what the appropriate mutational target size for common diseases should be. Thus, I consider a variety of models examining different parts of this parameter space. Some of these models cannot be rejected on the basis of existing exome sequencing and GWAS data. Other models may be more compatible with GWAS data if imperfect LD between causal variants and genotyped variants was properly taken into account. Overall, these models provide a framework consistent with existing empirical data with which to investigate the effect of recent population history and the genetic architecture of complex traits.
Recent growth and the heritability of complex traits
Using the models of demography, selection, and genetic architecture described above, I first examine the effect that population history has on the heritability of the trait. I find that population history has little effect on the heritability of the trait (Fig. S1), regardless of the values of , , and M used in the simulations. This is evidenced by the fact that in all three demographic scenarios investigated, the actual heritability estimated from each simulation replicate is close to , the value set in a constant size population. To further investigate the effect of recent growth on heritability, I divide the causal variants segregating at the end of the simulation into three categories. The first category consists of those SNPs that arose either further back in time than, or during the population bottleneck (“Before bottleneck” in Fig. 3). These mutations occurred >1960 generations ago. The second category consists of SNPs that arose after the population had recovered from the bottleneck, but further back in time than the recent population growth (“After bottleneck” in Fig. 3). These mutations arose between 1960 and 80 generations ago. The final category consists of SNPs that arose within the last 80 generations (“After growth” in Fig. 3). In the BN+growth model, these are the mutations that arose after the population expansion. Fig. 3A shows that the average heritability accounted for by mutations that arose at these three different time points is similar in both the BN+growth population (green boxes), and the BN population (orange boxes). Interestingly, when a mutation's effect on fitness is correlated with its effect on the trait (), mutations that arose in the last 80 generations, as a class, account for the greatest amount of the heritability (Fig. 3A).

Next, I investigate whether other features of genetic variation that affect the heritability (e.g. number of SNPs, mean allele frequency, mean effect size) are affected by recent population history. I find that recent growth has had little effect on the number of mutations that arose prior to the population bottleneck and are still segregating in the sample, the mean allele frequency, and the effect sizes of such mutations (Fig. 3B, Fig. 3C, and Fig. 3D). However, there is a different pattern for mutations that arose after the bottleneck, but more than 80 generations ago (those in the “After bottleneck” category). Recent population growth increases the number of such mutations (roughly 2-fold) relative to that found in the population that did not expand (Fig. 3B). Further, these mutations tend to be at lower frequency in the BN+growth population compared to the BN population (Fig. 3C). The only difference between the two models of population history on variants that arose during this time period is that genetic drift is weaker in the BN+growth population, compared to the BN population. Thus, fewer weakly deleterious mutations are lost from the BN+growth population, generating the pattern seen in Fig. 3B. The mutations that are not lost from the population tended to be at lower frequency in the larger population because they also are less likely to drift to higher frequency in the expanded population as compared to the non-expanded population. The mutations that arose within the last 80 generations also are affected by recent population history (those in the “After growth” category). As expected, recent population growth leads to a dramatic increase in the number of such SNPs (Fig. 3B). Further, the new mutations tend to be at lower frequency in the BN+growth population than in the BN population (Fig. 3C). More surprisingly, these SNPs tend to have weaker effect sizes on the trait in the BN+growth population than in the BN population (Fig. 3D). This observation can be explained by selection more effectively removing moderately and strongly deleterious mutations from the recently expanded population than from the non-recently expanded populations [65]. Thus, while recent population history affects the number of mutations, frequencies, and effect sizes of these mutations, it does so in such a way that the overall heritability of the trait appears unaffected by population history. Fig. S5 shows similar plots for the model where a mutation's effect on the trait is not correlated with its effect on fitness (τ = 0).
Recent growth increases the contribution of rare variants to the additive genetic variance
While population history does not affect the overall heritability of the trait, it can have a profound impact on the additive genetic variance (VA), and consequently, the heritability, attributable to low-frequency vs. common variants (Fig. 4). When a mutation's effect on fitness is correlated with its effect on the trait (τ = 0.5), more than 50% of the additive genetic variance in the trait is attributable to SNPs with frequency <0.5% in the population under all demographic scenarios, consistent with previous work showing of the importance of low-frequency variants [59], [72], [73]. Crucially, the amount of the variance attributable to rare variants (<0.1%) varies greatly due to demographic history. Roughly twice as much of the genetic variance of the trait in the recently expanded population (BN+ growth; green line in Fig. 4A) is accounted for by SNPs with frequency <0.05% than in the population that underwent the bottleneck, but did not expand (BN; orange line in Fig. 4A). The population that underwent ancient growth falls intermediate to the other two cases (Old growth; purple line in Fig. 4A). Similar results hold for other heritabilities and mutational targets (Fig. S6A–C). The situation is dramatically different if a SNP's effect on the trait is uncorrelated with its effect on fitness (τ = 0; Fig. 4B). Here little of the variance of the trait is accounted for by low-frequency variants, as seen by Eyre-Walker [59]. Additionally, under this model, demographic history does not make as substantial an impact on the amount of the additive genetic variance explained by SNPs at different frequencies, as suggested by Simons [38]. Again, similar results hold for other heritabilities considered and mutational target sizes (Fig. S6D–F). Thus, in some instances, recent population growth can result in a substantial increase in the amount of the genetic variance attributable to rare variants.
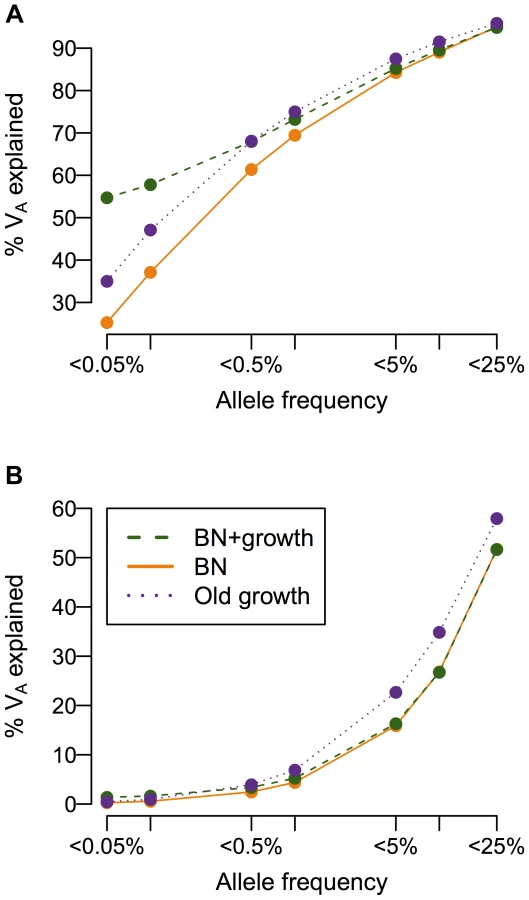
Recent growth increases genetic heterogeneity of disease
Population history also has a profound impact on the number of causal mutations in a sample of 1000 individuals who have been selected from the upper 40th percentile of the distribution of the quantitative trait (Fig. 5). These individuals can be thought of as cases. Here recent growth (BN+growth) is predicted to result in a substantial increase in the number of causal mutations compared to a population that has not undergone such recent growth (BN; orange vs. green boxes in Fig. 5A). In fact, a sample of 1000 cases from the BN+growth population is predicted to have nearly twice as many distinct causal mutations as a sample from the BN population. An explanation for these patterns is that many new deleterious causal mutations have arisen after the population has expanded in size. Because they are new and rare, they are only found in a small number of individuals. As such, each individual has his/her own set of low-frequency risk mutations. When aggregating this number across thousands of individuals, the total number of causal mutations in the sample from the BN+growth population is higher than in the BN population. Interestingly, the number of distinct causal mutations is actually higher in the BN+growth population than in the Old growth population (purple box in Fig. 5A).
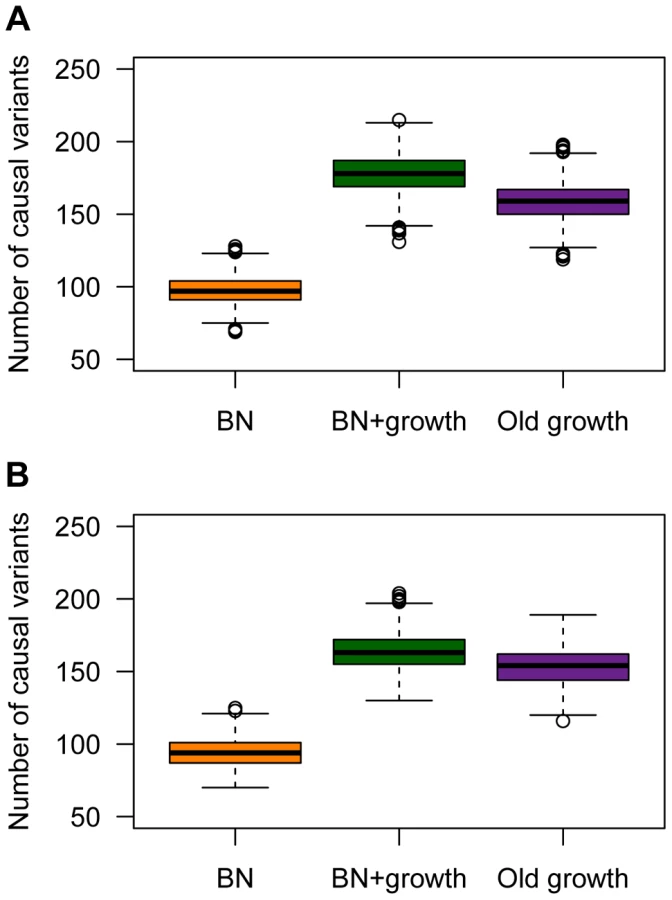
A similar increase in the number of causal variants in the sample from the recently expanded population relative to a non-expanded population is seen even when a SNP's effect on the trait is uncorrelated with its effect on fitness (τ = 0; Fig. 5B). This pattern is due to the fact that there is the same number of rare causal variants in the BN+growth population even when τ = 0. However, when τ = 0, many of these rare causal mutations have smaller effect sizes, and consequently, do not account for much of the phenotypic variance of the trait.
To further explore this issue, I examine how much of the phenotypic variance (VP) in the population can be accounted for by the SNPs that explain the most VP (Fig. 6). When τ = 0.5, the top SNPs that explain most of the variance account for less of it in the population that recently expanded (BN+growth) than in the population that did not (BN). For example, when , the 25 SNPs that account for the most VP will account for 5% of the VP in the BN population (orange line in Fig. 6A). In contrast, for the BN+growth population (green line in Fig. 6A), the 25 SNPs that account for the most VP will only explain <3.5% of it. Put another way, the top 25 SNPs that explain the most variance account for >90% of the VA contained within the mutational target in the BN population, but <70% of the VA in the BN+growth population. These results suggest that, if mutational effects on disease are correlated with their effects on fitness, many of the additional rare causal variants found in a recently expanded population, may, in aggregate, explain a substantial proportion (say 20%) of the heritability of the trait.

If a mutation's effect on fitness is independent of its effect on disease (τ = 0), then the top SNPs that explain the most variance account for almost all of the VA contained within the mutational target (Fig. 6B). For example, in the model where , the top 25 SNPs will account for nearly 5% of the VP, regardless of the demographic history of the population. Put another way, here the top 25 SNPs account for the majority of the VA that is contained within the mutational target, and this pattern is not affected by the demography of the population. This finding supports the previous statement that, if a mutation's effect on fitness is independent of its effect on the trait, many of the extra causal mutations seen in Fig. 5B in the recently expanded population actually contribute very little to the overall VP of the trait. Similar results are found for other values of and mutational target sizes (Fig. S7).
Effect of demography on the power of association tests
Next, I investigate how different demographic histories affect the power to associate SNPs with a trait in a sample of 1000 cases and 1000 controls. Most power simulations for association tests examine the power to detect a given causal variant conditional on its allele frequency and/or effect size. Using this approach, I find that power to detect the SNPs that explain the greatest amount of VA is actually higher in the population that recently expanded (BN+growth) than in the population that only underwent a bottleneck (BN; Text S2). However, recent population growth has a more limited effect on the power to detect a given association when conditioning on the allele frequency or odds ratio of the causal SNP (Text S2).
The power analyses summarized above refer to the power to detect a given causal variant, conditional on various attributes of it. However, the number of causal variants, their frequencies, and their effect sizes are random variables that are influenced by the evolutionary process experienced by the population under study. Thus, it is also useful to examine the expected number of causal SNPs with P-values less than the significance threshold across the different models of demographic history (Fig. 7). The expected number of causal SNPs detected in a study of a given sample size will account for both the power to detect a given variant as well as the number, frequency distribution, and effect size distribution of causal variants in the population. It also directly answers the relevant question for researchers who are planning and interpreting an association study: Under a given model of genetic architecture, with a given sample size, how many significant associations would I expect to detect?
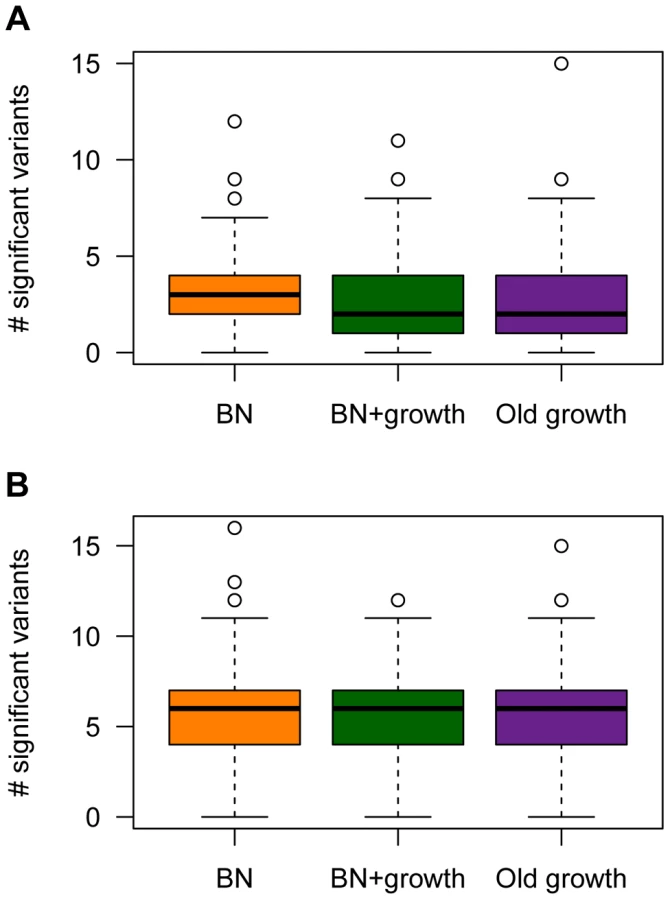
Under the model where a mutation's effect on fitness is correlated with its effect on the trait (τ = 0.5), , and M = 70 kb, fewer causal mutations are detected in the populations that have undergone ancient (Old growth; purple box in Fig. 7A) or recent (BN+growth; green box in Fig. 7A) expansions, relative to the population that only underwent a recent bottleneck (BN; orange box in Fig. 7A). Similar trends are seen for the other models of and M (Fig. S4). However, when , sample sizes of 1000 cases and controls are too small to detect almost any associations with P<1×10−5, regardless of the demography of the population. When using a less stringent significance threshold (P<0.01), , and M = 70 kb, a median of 10 causal loci are associated with the trait in the BN population (Fig. S8A). However, a median of only 8 causal loci are detected in the BN+growth population. Again, similar trends are noted for the other models of and M (Fig. S8). However, when , a median of 2 causal SNPs are detected at P<0.01 for all three demographic models. This result is due to the very low power to detect an association for causal variants with very small effect sizes using samples of 1000 cases and 1000 controls, regardless of the demography of the population. Nevertheless, even here, a higher proportion of simulation replicates have detected at least 3 associations in the BN population (54%) than in the BN+growth population (41%). Taken together, this analysis suggests that recent population growth can result in a decrease in the expected number of associations to be detected in a given sample size. Thus, while recent growth may increase power to detect the SNP that explains the greatest amount of the variance, and have little effect on power to detect a given SNP conditional on its frequency or effect size, it enriches the frequency distribution for rare causal variants. The power to detect such variants using single-marker association tests is low, decreasing the expected number of significant associations to be detected in the population that recently expanded.
However, demographic history has no clear effect on the number of causal loci detected with a given sample size when the mutation's effect on fitness is independent of its effect on the trait (τ = 0; Fig. 7B, Fig. S4D–F, and Fig. S8E–H). For some models, the BN+growth population appears to have a higher number of significant associations than in the BN population (Fig. S4D and Fig. S4E). However, this pattern is not consistently seen across models or significance thresholds. Similarly, when using a significance threshold of P<0.01, the Old growth population appears to show a greater number of significant association (Fig. S8) than either of the other two models of population history. This pattern may be due to the slight, but noticeable, increase in the for the Old growth population (Fig. S1).
Researchers have suggested that the amount of the additive genetic variance (VA) explained by a set of SNPs increases when the stringency of the P-value threshold for including SNPs in the set is decreased [74]–[76]. Fig. 8 shows the amount of VA explained by SNPs having single-marker P-values less than the threshold specified on the x-axis for the model where and M = 70 kb. When a SNP's effect on the trait is correlated with its effect on fitness (τ = 0.5), population history has an important effect on the amount of VA accounted for by SNPs with P-value less than a given threshold (Fig. 8A). Specifically, recent population growth decreases the amount of VA accounted for by SNPs at all P-value thresholds, relative to what is seen in a population that has not expanded. For example, SNPs having P<0.05 account for about 30% of the VA in the BN population. SNPs with P<0.05 account for only 20% of VA in the BN+growth population. Including all SNPs detected in the case-control study captures only 70% of the VA contained within the mutational target in the BN+growth population. The reason for this is that many of the rare variants that account for the VA of the trait in the population are not present in the sample of 1000 cases and 1000 controls. When τ = 0, population history has little affect on the amount of VA accounted for by SNPs with a given P-value (Fig. 8B). Including all SNPs present in the association study captures over 95% of the VA contained within the mutational target. This finding is not surprising in light of the observation (Fig. 4B) that much of the VA is accounted for by common variants when τ = 0, and such variants are likely to be present in the sample of 1000 cases and 1000 controls. Qualitatively similar trends are seen for other heritablities and mutational target sizes (Fig. S9).

Discussion
I have shown that very recent population growth can have an impact on patterns of deleterious genetic variation and the genetic architecture of complex traits. Specifically, I show that recent population growth leads to an increase in the proportion of nonsynonymous SNPs relative to that expected in non-expanded populations. Further, this recent growth is predicted to have affected the genetic architecture of some complex traits. This result has implications for discovering the “missing heritability” in different human populations and detecting causal variants that may also affect reproductive fitness.
While I have shown that demographic history greatly affects the proportion and frequencies of deleterious mutations segregating in the population, it is interesting that demography does not have a large effect on the overall genetic load of the population. Haldane has shown that the genetic load at equilibrium contributed by a particular mutation is independent of the strength of selection acting on the particular mutation and the frequency of that mutation [66]. Strongly deleterious mutations will have a large effect on fitness, but will be kept at lower frequency by negative selection than mutations of weaker effect. Weakly deleterious mutations will not have as large of an effect, but they can be at higher frequency in the population. Haldane suggested that these effects should cancel each other out. Haldane's result was derived for a simple model with a constant population size. It was unclear whether this result would hold when considering populations with bottlenecks and recent growth, though previous theoretical results suggested that smaller populations may be expected to have a higher load [77]. Here I have shown that Haldane's result applies under certain complex demographic models. Further work is required to determine whether this trend holds in other species and demographic models, and whether this trend holds for models involving dominance.
It is also important to appreciate that while my analysis as well as that of Simons et al. [38] show that recent population history has not affected the overall genetic load of the population, these results should not be taken to imply that population history has had no effect on patterns of deleterious variation. First, the way in which the populations arrive at their load differs across populations with different histories. Populations that have recently expanded have more of the load accounted for by rare variants than populations that have not recently expanded. Further, genetic load is one specific feature of deleterious genetic variation. Other statistics, like the proportion of nonsynonymous SNPs, are very sensitive to differences in population history. While it has been shown that differences in population history between European and African populations have affected the proportion of nonsynonymous SNPs in the two populations [54], here I demonstrate that the influence of population history on the proportion of nonsynonymous SNPs is predicted to apply on a much more recent timescale, and to populations that are much more similar to each other than Europeans and Africans. In sum, taken together, these results suggest that population history affects patterns of deleterious genetic variation found in different populations.
I find that population history is predicted to have little effect on the overall amount of additive genetic variance for a trait seen in different populations. As such, assuming a common environmental variance across populations, the heritability of a trait is predicted to be similar across populations. This finding suggests that if differences in the heritability of a trait are detected across populations, these differences are more likely to be due to differing environmental effects, rather than due to different amounts of additive genetic variance. For example, it has been suggested that the heritability of height in a West African population is less than that typically estimated from European populations [78]. My results would argue that such a difference would be due to shifts in the environmental variance, rather than changes in amount of additive genetic variance as a result of differences in recent population history.
A major conclusion of this study is that recent population growth has a greater effect on the architecture of traits when a mutation's effect on fitness is correlated with its effect on the phenotype than when the mutation's effect on fitness is independent of its effect on the phenotype. The extent to which mutations that increase risk of disease are under negative selection remains unclear. While it is intuitive that disease mutations should be under negative selection, most common diseases have an onset after reproductive age, and as such, may not be correlated with reproductive fitness. However, it is possible that mutations increasing risk to late-onset disease may have pleiotropic effects and could affect traits related to reproduction [59], [79]. In fact, it has been suggested that pleiotropy is rather frequent for many common SNPs associated with disease [80], and may apply to rare variants as well. Additionally, genes and common variants associated with common diseases show signatures of negative selection [81], [82], suggesting that disease variants may be under negative selection. A third line of evidence comes from studies of model organisms. A mutagenesis study [83] found that P-element insertions that contributed the most to the variance in bristle number in Drosophila tended to reduce viability. A fourth line of evidence suggesting that a mutation's effect on complex disease may be correlated with fitness comes from an empirical analysis of GWAS data. Looking at over 350 susceptibility SNPs across eight categories of phenotypes, Park et al. [84] found that low-frequency SNPs tended to have larger effect sizes than more common SNPs (significantly so for type 1 diabetes, height, and LDL levels), even after correcting for the ascertainment bias resulting from the reduced power to detect associations with low-frequency SNPs of weak effect. Such a correlation is expected under models where a mutation's effect on fitness is correlated with its effect on the trait (τ = 0.5; Fig. S10A). However, a correlation between a mutation's effect on disease and its frequency is not expected under a model where a mutation's effect on disease is independent of its effect on fitness (τ = 0; Fig. S10B). There is little direct evidence to indicate whether this result holds for low-frequency variants in coding regions. But, selection is likely to be stronger (per base pair) in coding regions than throughout noncoding regions of the genome [85], suggesting this result should hold for coding regions as well. Finally, the correlation between a mutation's effect on fitness and its effect on a trait is likely to depend on the particular trait involved. While further empirical and theoretical work is needed in this area, all of these lines of evidence suggest that, for some traits, it is plausible that a mutation's effect on fitness could indeed be correlated with its effect on disease.
Further rationale for considering models where a mutation's effect on fitness is correlated with its effect on the trait comes from exome sequencing studies themselves. A major assumption made in exome sequencing studies is that some of the missing heritability can be explained by rare variants of large effect that increase risk to disease [3], [4]. If there is no correlation between a mutation's effect on disease and its effect on fitness, then there is no reason for the effects on disease risk to be greater for rare variants than for more common variants. Under this model (where fitness effects are independent of trait effects), effect sizes would be randomly assigned to SNPs, regardless of their allele frequency. On the other hand, if a mutation's effect on fitness is correlated with its effect on disease, then the SNPs with the strongest effects on disease are likely to be the most deleterious ones. As such, they will also be the most rare in the population due to negative selection. Because the exome sequencing paradigm essentially assumes that the effect of a coding region mutation on disease is correlated with its effect on fitness, it is important to investigate the proprieties of such a model under different population histories.
My models make several predictions that can be tested with empirical data. While these models have been developed to apply to exome sequencing data, because the predictions are robust to the mutational target size and heritability accounted for by the mutations in the target region (Fig. S1, Fig. S4, Fig. S6–Fig. S9), they should apply to GWAS data as well, especially if low-frequency variants are imputed from a reference panel, like the 1000 Genomes Project. First, the models predict that if a mutation's effect on fitness is correlated with its effect on the trait, common variants should account for more of the heritability in a population that did not expand than in one that had recently expanded. This prediction can be tested by analyzing GWAS data in expanded vs. non-expanded populations. Second, for a given sample size, if a mutation's effect on fitness is correlated with its effect on the trait, the models predict that fewer significant associations will be detected in the recently expanded population than in a population that has not expanded. This prediction can also be tested by comparing the number of significant associations detected in GWAS data from the expanded population vs. the non-expanded population. Third, the prediction that, if a mutation's effect on fitness is correlated with its effect on the trait, low-frequency variants should account for more of the heritability in the recently expanded population than in a non-expanded population can be tested directly once large-scale exome sequencing data in both expanded and non-expanded populations has been collected. Failing to find these patterns in GWAS and exome sequencing data would suggest that there is little correlation between a mutation's effect on fitness and its effect on the trait.
Several recent studies have used results from population genetic models to guide the design and interpretation of association studies of rare variants [86], [87]. My results are especially complementary to those Zuk et al. [87]. In particular, they argue that a population expansion does not increase the proportion of the disease due to new alleles. My finding of a similar contribution of young alleles to the heritability in expanded vs. non-expanded populations (Fig. 3A) supports their conclusion, despite different modeling assumptions in the two studies. Zuk et al. [87] also argue that the “role” of rare variants in disease is increased in an expanding population. Here I provide a more detailed analysis of this topic using an explicit quantitative genetic model and evaluate the conditions under which recent population growth accentuates the contribution of rare variants to the heritability of the trait.
One important limitation of the present study is that I evaluated the power of single-marker association tests, rather than gene-based association tests. It has been suggested that single-marker tests may be under-powered relative to gene-based tests for detecting associations with rare variants [37], [88]. I did not consider gene-based association tests in the present study because the present simulations assume that all SNPs are independent of each other. Thus, there is no way to simulate genes containing multiple SNPs having the appropriate LD structure in this framework. Future work could simulate larger genic regions using other approaches [56]. However, the analysis of the performance of single-marker association tests still provides important insights for understanding how demography affects the ability to map genes for complex traits. First, many association studies of rare variants include single-marker association tests, even if they also consider gene-based tests [23]–[25], [88]. Thus, my results are directly applicable to designing and interpreting such studies. Second, several gene-based association tests combine the single-marker association tests in various ways. Thus, the manner in which single-marker signals are affected by demography is relevant for such tests. Third, it is not clear that gene-based tests are always superior to single-marker tests. A recent study [89] has suggested that gene-based tests based on single-marker association statistics may be more powerful than other “burden tests.” Further, performance of gene-based association tests is known to decrease if many non-causal SNPs are included [22], [87], [89]. Also, if causal variants are scattered across many distinct genes, then gene-based association tests may not provide an increase in power over single-marker tests [30]. As sample sizes continue to grow, it is likely that single-marker tests will be more frequently used for sequencing-based association studies because they are simpler to implement and eliminate the need to decide how to combine variants within a gene. Thus, insights gained from the analysis of single-marker association tests in the present study should still be useful.
Another possible limitation of this study is that my models do not allow some mutations affecting complex traits to be beneficial or under balancing selection. There is some evidence that loci associated with certain traits (obesity and type 2 diabetes, in particular [90], [91]) may have been affected by positive selection. However, this pattern was found for common variants outside of coding regions. It is not clear how many nonsynonymous causal mutations would be expected to be under positive selection. Additionally, loci under positive or balancing selection should have already been detected in GWAS (because they would be common variants), and are not the type of loci researchers are aiming to discover through exome sequencing studies.
My results suggest that if a mutation's effect on fitness is correlated with its effect on the trait, recent population history can have an important effect on the ability to detect associations with causal variants. In the simulations, fewer causal SNPs were significantly associated with disease in the population that experienced recent growth as compared to the population that did not expand. This result would imply that, in order to detect the greatest number of causal loci for a given sample size, it would be better to focus on populations that experienced a bottleneck, but did not experience a recent population expansion. Currently, it is not clear which populations meet this criterion. Further sequencing of large samples of individuals will be required to determine which populations have not experienced recent population growth. For a variety of reasons, most GWAS have been done in large samples of cases and controls of European ancestry [92]. The same trend may hold for exome and genome re-sequencing studies. Recent genetic studies, as well as historical records indicate that many European populations are precisely those that experienced the type of extreme, recent population growth simulated in this study [36]. The simulations presented here suggest that focusing on such populations may not discover the largest number of causal variants for a given number of individuals sequenced.
An important goal in human genetics is to understand disease risk in populations throughout the globe. While focusing on populations that have not experienced ancient or recent growth may yield the largest list of putative causal loci, it is currently not clear whether such an approach will lead to increased understanding of the genetic basis of disease in all populations—not just the population under study. Analyses of common variants implicated through GWAS suggest that loci associated with traits in European populations also affect the traits in other non-European populations [93]–[96]. However, it is unclear whether this trend also holds for rare variants that have arisen after the populations have split from each other. Thus, in order to understand the genetic basis of complex traits across the globe, it will be important to study populations that have recently expanded in addition to those more stable in size, recognizing that larger sample sizes in populations that have recently expanded will be necessary to achieve comparable power to that in non-expanded populations.
Finally, these results are directly relevant for finding the “missing heritability” in different populations. If a mutation's effect on disease is correlated with its effect on fitness, then more of the heritability will be explained by very rare variants in a population that experienced a recent expansion than in a population that did not recently expand. Additionally, the variants detected by single-marker association tests explain less of the heritability in a recently expanded population than in a population that did not recently expand. Thus, while the overall heritability of a trait may not be variable across populations, our ability to discover the variants that account for it is likely to vary across populations due to differences in demographic history.
Supporting Information
Zdroje
1. AltshulerD, DalyMJ, LanderES (2008) Genetic mapping in human disease. Science 322 : 881–888.
2. StrangerBE, StahlEA, RajT (2011) Progress and promise of genome-wide association studies for human complex trait genetics. Genetics 187 : 367–383.
3. ManolioTA, CollinsFS, CoxNJ, GoldsteinDB, HindorffLA, et al. (2009) Finding the missing heritability of complex diseases. Nature 461 : 747–753.
4. GibsonG (2012) Rare and common variants: Twenty arguments. Nat Rev Genet 13 : 135–145.
5. EE, FlintJ, GibsonG, KongA, LealSM, et al. (2010) Missing heritability and strategies for finding the underlying causes of complex disease. Nat Rev Genet 11 : 446–450.
6. NgSB, TurnerEH, RobertsonPD, FlygareSD, BighamAW, et al. (2009) Targeted capture and massively parallel sequencing of 12 human exomes. Nature 461 : 272–276.
7. ShendureJ, JiH (2008) Next-generation DNA sequencing. Nat Biotechnol 26 : 1135–1145.
8. WuMC, LeeS, CaiT, LiY, BoehnkeM, et al. (2011) Rare-variant association testing for sequencing data with the sequence kernel association test. Am J Hum Genet 89 : 82–93.
9. MadsenBE, BrowningSR (2009) A groupwise association test for rare mutations using a weighted sum statistic. PLoS Genet 5: e1000384.
10. B, LealSM (2008) Methods for detecting associations with rare variants for common diseases: Application to analysis of sequence data. Am J Hum Genet 83 : 311–321.
11. LiB, LealSM (2009) Discovery of rare variants via sequencing: Implications for the design of complex trait association studies. PLoS Genet 5: e1000481.
12. BamshadMJ, NgSB, BighamAW, TaborHK, EmondMJ, et al. (2011) Exome sequencing as a tool for Mendelian disease gene discovery. Nat Rev Genet 12 : 745–755.
13. BamshadMJ, ShendureJA, ValleD, HamoshA, LupskiJR, et al. (2012) The centers for Mendelian genomics: A new large-scale initiative to identify the genes underlying rare Mendelian conditions. Am J Med Genet A 158A: 1523–1525.
14. NgSB, BighamAW, BuckinghamKJ, HannibalMC, McMillinMJ, et al. (2010) Exome sequencing identifies MLL2 mutations as a cause of Kabuki syndrome. Nat Genet 42 : 790–793.
15. NgSB, BuckinghamKJ, LeeC, BighamAW, TaborHK, et al. (2010) Exome sequencing identifies the cause of a Mendelian disorder. Nat Genet 42 : 30–35.
16. NgSB, NickersonDA, BamshadMJ, ShendureJ (2010) Massively parallel sequencing and rare disease. Hum Mol Genet 19: R119–24.
17. 1000 Genomes Project Consortium, Durbin RM, Abecasis GR, Altshuler DL, Auton A, et al (2010) A map of human genome variation from population-scale sequencing. Nature 467 : 1061–1073.
18. 1000 Genomes Project Consortium, Abecasis GR, Auton A, Brooks LD, DePristo MA, et al (2012) An integrated map of genetic variation from 1,092 human genomes. Nature 491 : 56–65.
19. CirulliET, GoldsteinDB (2010) Uncovering the roles of rare variants in common disease through whole-genome sequencing. Nat Rev Genet 11 : 415–425.
20. GorlovIP, GorlovaOY, SunyaevSR, SpitzMR, AmosCI (2008) Shifting paradigm of association studies: Value of rare single-nucleotide polymorphisms. Am J Hum Genet 82 : 100–112.
21. SchorkNJ, MurraySS, FrazerKA, TopolEJ (2009) Common vs. rare allele hypotheses for complex diseases. Curr Opin Genet Dev 19 : 212–219.
22. KiezunA, GarimellaK, DoR, StitzielNO, NealeBM, et al. (2012) Exome sequencing and the genetic basis of complex traits. Nat Genet 44 : 623–630.
23. HelgasonH, SulemP, DuvvariMR, LuoH, ThorleifssonG, et al. (2013) A rare nonsynonymous sequence variant in C3 is associated with high risk of age-related macular degeneration. Nat Genet 45 : 1371–1374.
24. SeddonJM, YuY, MillerEC, ReynoldsR, TanPL, et al. (2013) Rare variants in CFI, C3 and C9 are associated with high risk of advanced age-related macular degeneration. Nat Genet 45 : 1366–1370.
25. ZhanX, LarsonDE, WangC, KoboldtDC, SergeevYV, et al. (2013) Identification of a rare coding variant in complement 3 associated with age-related macular degeneration. Nat Genet 45 : 1375–1379.
26. HeinzenEL, DepondtC, CavalleriGL, RuzzoEK, WalleyNM, et al. (2012) Exome sequencing followed by large-scale genotyping fails to identify single rare variants of large effect in idiopathic generalized epilepsy. Am J Hum Genet 91 : 293–302.
27. NeedAC, McEvoyJP, GennarelliM, HeinzenEL, GeD, et al. (2012) Exome sequencing followed by large-scale genotyping suggests a limited role for moderately rare risk factors of strong effect in schizophrenia. Am J Hum Genet 91 : 303–312.
28. HuntKA, MistryV, BockettNA, AhmadT, BanM, et al. (2013) Negligible impact of rare autoimmune-locus coding-region variants on missing heritability. Nature 498 : 232–235.
29. LiuL, SaboA, NealeBM, NagaswamyU, StevensC, et al. (2013) Analysis of rare, exonic variation amongst subjects with autism spectrum disorders and population controls. PLoS Genet 9: e1003443.
30. LohmuellerKE, SparsoT, LiQ, AnderssonE, KorneliussenT, et al. (2013) Whole-exome sequencing of 2,000 Danish individuals and the role of rare coding variants in type 2 diabetes. Am J Hum Genet 93 : 1072–1086.
31. TennessenJA, BighamAW, O'ConnorTD, FuW, KennyEE, et al. (2012) Evolution and functional impact of rare coding variation from deep sequencing of human exomes. Science 337 : 64–69.
32. NelsonMR, WegmannD, EhmMG, KessnerD, St JeanP, et al. (2012) An abundance of rare functional variants in 202 drug target genes sequenced in 14,002 people. Science 337 : 100–104.
33. FuW, O'ConnorTD, JunG, KangHM, AbecasisG, et al. (2013) Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants. Nature 493 : 216–220.
34. MarthGT, YuF, IndapAR, GarimellaK, GravelS, et al. (2011) The functional spectrum of low-frequency coding variation. Genome Biol 12: R84–2011-12-9-r84.
35. CoventryA, Bull-OttersonLM, LiuX, ClarkAG, MaxwellTJ, et al. (2010) Deep resequencing reveals excess rare recent variants consistent with explosive population growth. Nat Commun 1 : 131.
36. KeinanA, ClarkAG (2012) Recent explosive human population growth has resulted in an excess of rare genetic variants. Science 336 : 740–743.
37. KryukovGV, ShpuntA, StamatoyannopoulosJA, SunyaevSR (2009) Power of deep, all-exon resequencing for discovery of human trait genes. Proc Natl Acad Sci U S A 106 : 3871–3876.
38. SimonsYB, TurchinMC, PritchardJK, SellaG (2014) The deleterious mutation load is insensitive to recent population history. Nat Genet 46 : 220–224.
39. WrightAF, CarothersAD, PirastuM (1999) Population choice in mapping genes for complex diseases. Nat Genet 23 : 397–404.
40. PritchardJK, PrzeworskiM (2001) Linkage disequilibrium in humans: Models and data. Am J Hum Genet 69 : 1–14.
41. PeltonenL, PalotieA, LangeK (2000) Use of population isolates for mapping complex traits. Nat Rev Genet 1 : 182–190.
42. Service S, DeYoung J, Karayiorgou M, Roos JL, Pretorious H, et al (2006) Magnitude and distribution of linkage disequilibrium in population isolates and implications for genome-wide association studies. Nat Genet 38 : 556–560.
43. ArdlieKG, KruglyakL, SeielstadM (2002) Patterns of linkage disequilibrium in the human genome. Nat Rev Genet 3 : 299–309.
44. ReichDE, CargillM, BolkS, IrelandJ, SabetiPC, et al. (2001) Linkage disequilibrium in the human genome. Nature 411 : 199–204.
45. AkeyJM, EberleMA, RiederMJ, CarlsonCS, ShriverMD, et al. (2004) Population history and natural selection shape patterns of genetic variation in 132 genes. PLoS Biol 2: e286.
46. BoykoAR, WilliamsonSH, IndapAR, DegenhardtJD, HernandezRD, et al. (2008) Assessing the evolutionary impact of amino acid mutations in the human genome. PLoS Genet 4: e1000083.
47. GutenkunstRN, HernandezRD, WilliamsonSH, BustamanteCD (2009) Inferring the joint demographic history of multiple populations from multidimensional SNP frequency data. PLoS Genet 5: e1000695.
48. KeinanA, MullikinJC, PattersonN, ReichD (2007) Measurement of the human allele frequency spectrum demonstrates greater genetic drift in East Asians than in Europeans. Nat Genet 39 : 1251–1255.
49. LohmuellerKE, BustamanteCD, ClarkAG (2009) Methods for human demographic inference using haplotype patterns from genomewide single-nucleotide polymorphism data. Genetics 182 : 217–231.
50. VoightBF, AdamsAM, FrisseLA, QianY, HudsonRR, et al. (2005) Interrogating multiple aspects of variation in a full resequencing data set to infer human population size changes. Proc Natl Acad Sci U S A 102 : 18508–18513.
51. TennessenJA, O'ConnorTD, BamshadMJ, AkeyJM (2011) The promise and limitations of population exomics for human evolution studies. Genome Biol 12 : 127.
52. GazaveE, MaL, ChangD, CoventryA, GaoF, et al. (2014) Neutral genomic regions refine models of recent rapid human population growth. Proc Natl Acad Sci U S A 111 : 757–762.
53. LohmuellerKE, BustamanteCD, ClarkAG (2010) The effect of recent admixture on inference of ancient human population history. Genetics 185 : 611–622.
54. LohmuellerKE, IndapAR, SchmidtS, BoykoAR, HernandezRD, et al. (2008) Proportionally more deleterious genetic variation in European than in African populations. Nature 451 : 994–997.
55. SawyerSA, HartlDL (1992) Population genetics of polymorphism and divergence. Genetics 132 : 1161–1176.
56. HernandezRD (2008) A flexible forward simulator for populations subject to selection and demography. Bioinformatics 24 : 2786–2787.
57. HoggartCJ, Chadeau-HyamM, ClarkTG, LamparielloR, WhittakerJC, et al. (2007) Sequence-level population simulations over large genomic regions. Genetics 177 : 1725–1731.
58. ClampM, FryB, KamalM, XieX, CuffJ, et al. (2007) Distinguishing protein-coding and noncoding genes in the human genome. Proc Natl Acad Sci U S A 104 : 19428–19433.
59. Eyre-WalkerA (2010) Evolution in health and medicine Sackler Colloquium: Genetic architecture of a complex trait and its implications for fitness and genome-wide association studies. Proc Natl Acad Sci U S A 107 Suppl 11752–1756.
60. VisscherPM, HillWG, WrayNR (2008) Heritability in the genomics era—concepts and misconceptions. Nat Rev Genet 9 : 255–266.
61. YangJ, LeeSH, GoddardME, VisscherPM (2011) GCTA: A tool for genome-wide complex trait analysis. Am J Hum Genet 88 : 76–82.
62. ZaitlenN, KraftP (2012) Heritability in the genome-wide association era. Hum Genet 131 : 1655–1664.
63. DempsterER, LernerIM (1950) Heritability of threshold characters. Genetics 35 : 212–236.
64. RischNJ (2000) Searching for genetic determinants in the new millennium. Nature 405 : 847–856.
65. GazaveE, ChangD, ClarkAG, KeinanA (2013) Population growth inflates the per-individual number of deleterious mutations and reduces their mean effect. Genetics 195 : 969–978.
66. HaldaneJBS (1937) The effect of variation on fitness. Am Nat 71 : 337–349.
67. MullerHJ (1950) Our load of mutations. Am J Hum Genet 2 : 111–176.
68. PurcellSM, MoranJL, FromerM, RuderferD, SolovieffN, et al. (2014) A polygenic burden of rare disruptive mutations in schizophrenia. Nature 506 : 185–190.
69. LangeLA, HuY, ZhangH, XueC, SchmidtEM, et al. (2014) Whole-exome sequencing identifies rare and low-frequency coding variants associated with LDL cholesterol. Am J Hum Genet 94 : 233–245.
70. WrayNR, PurcellSM, VisscherPM (2011) Synthetic associations created by rare variants do not explain most GWAS results. PLoS Biol 9: e1000579.
71. VisscherPM, BrownMA, McCarthyMI, YangJ (2012) Five years of GWAS discovery. Am J Hum Genet 90 : 7–24.
72. PritchardJK (2001) Are rare variants responsible for susceptibility to complex diseases? Am J Hum Genet 69 : 124–137.
73. ThorntonKR, ForanAJ, LongAD (2013) Properties and modeling of GWAS when complex disease risk is due to non-complementing, deleterious mutations in genes of large effect. PLoS Genet 9: e1003258.
74. International Schizophrenia Consortium (2009) PurcellSM, WrayNR, StoneJL, VisscherPM, et al. (2009) Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 460 : 748–752.
75. Lango AllenH, EstradaK, LettreG, BerndtSI, WeedonMN, et al. (2010) Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature 467 : 832–838.
76. WilliamsSM, HainesJL (2011) Correcting away the hidden heritability. Ann Hum Genet 75 : 348–350.
77. KimuraM, MaruyamaT, CrowJF (1963) The mutation load in small populations. Genetics 48 : 1303–1312.
78. RobertsDF, BillewiczWZ, McGregorIA (1978) Heritability of stature in a West African population. Ann Hum Genet 42 : 15–24.
79. WrightA, CharlesworthB, RudanI, CarothersA, CampbellH (2003) A polygenic basis for late-onset disease. Trends Genet 19 : 97–106.
80. SivakumaranS, AgakovF, TheodoratouE, PrendergastJG, ZgagaL, et al. (2011) Abundant pleiotropy in human complex diseases and traits. Am J Hum Genet 89 : 607–618.
81. MaherMC, UricchioLH, TorgersonDG, HernandezRD (2012) Population genetics of rare variants and complex diseases. Hum Hered 74 : 118–128.
82. PavardS, MetcalfCJ (2007) Negative selection on BRCA1 susceptibility alleles sheds light on the population genetics of late-onset diseases and aging theory. PLoS One 2: e1206.
83. LymanRF, LawrenceF, NuzhdinSV, MackayTF (1996) Effects of single P-element insertions on bristle number and viability in Drosophila melanogaster. Genetics 143 : 277–292.
84. ParkJH, GailMH, WeinbergCR, CarrollRJ, ChungCC, et al. (2011) Distribution of allele frequencies and effect sizes and their interrelationships for common genetic susceptibility variants. Proc Natl Acad Sci U S A 108 : 18026–18031.
85. TorgersonDG, BoykoAR, HernandezRD, IndapA, HuX, et al. (2009) Evolutionary processes acting on candidate cis-regulatory regions in humans inferred from patterns of polymorphism and divergence. PLoS Genet 5: e1000592.
86. Agarwala V, Flannick J, Sunyaev S, GoT2D Consortium, Altshuler D (2013) Evaluating empirical bounds on complex disease genetic architecture. Nat Genet 45 : 1418–1427.
87. ZukO, SchaffnerSF, SamochaK, DoR, HechterE, et al. (2014) Searching for missing heritability: Designing rare variant association studies. Proc Natl Acad Sci U S A 111: E455–64.
88. DoR, KathiresanS, AbecasisGR (2012) Exome sequencing and complex disease: Practical aspects of rare variant association studies. Hum Mol Genet 21: R1–9.
89. KinnamonDD, HershbergerRE, MartinER (2012) Reconsidering association testing methods using single-variant test statistics as alternatives to pooling tests for sequence data with rare variants. PLoS One 7: e30238.
90. CastoAM, FeldmanMW (2011) Genome-wide association study SNPs in the human genome diversity project populations: Does selection affect unlinked SNPs with shared trait associations? PLoS Genet 7: e1001266.
91. ChenR, CoronaE, SikoraM, DudleyJT, MorganAA, et al. (2012) Type 2 diabetes risk alleles demonstrate extreme directional differentiation among human populations, compared to other diseases. PLoS Genet 8: e1002621.
92. BustamanteCD, BurchardEG, De la VegaFM (2011) Genomics for the world. Nature 475 : 163–165.
93. CarlsonCS, MatiseTC, NorthKE, HaimanCA, FesinmeyerMD, et al. (2013) Generalization and dilution of association results from European GWAS in populations of non-European ancestry: The PAGE study. PLoS Biol 11: e1001661.
94. ShrinerD, AdeyemoA, GerryNP, HerbertA, ChenG, et al. (2009) Transferability and fine-mapping of genome-wide associated loci for adult height across human populations. PLoS One 4: e8398.
95. WatersKM, StramDO, HassaneinMT, Le MarchandL, WilkensLR, et al. (2010) Consistent association of type 2 diabetes risk variants found in Europeans in diverse racial and ethnic groups. PLoS Genet 6: e1001078.
96. MarigortaUM, NavarroA (2013) High trans-ethnic replicability of GWAS results implies common causal variants. PLoS Genet 9: e1003566.
Štítky
Genetika Reprodukčná medicínaČlánok vyšiel v časopise
PLOS Genetics
2014 Číslo 5
- Gynekologové a odborníci na reprodukční medicínu se sejdou na prvním virtuálním summitu
- Je „freeze-all“ pro všechny? Odborníci na fertilitu diskutovali na virtuálním summitu
Najčítanejšie v tomto čísle
- PINK1-Parkin Pathway Activity Is Regulated by Degradation of PINK1 in the Mitochondrial Matrix
- Null Mutation in PGAP1 Impairing Gpi-Anchor Maturation in Patients with Intellectual Disability and Encephalopathy
- Phosphorylation of a WRKY Transcription Factor by MAPKs Is Required for Pollen Development and Function in
- p53 Requires the Stress Sensor USF1 to Direct Appropriate Cell Fate Decision